НОВЫЙ СЛУЧАЙ РЕКОМБИНАЦИИ МЕЖДУ ЯДЕРНЫМ - И МИТОХОНДРИАЛЬНЫМ ГЕНОМАМИ В РОДЕ Calliope Gould, 1836 (Muscicapidae, Aves): ГИПОТЕЗА ПРОИСХОЖДЕНИЯ
←
→
Транскрипция содержимого страницы
Если ваш браузер не отображает страницу правильно, пожалуйста, читайте содержимое страницы ниже
ГЕНЕТИКА, 2019, том 55, № 1, с. 81–93
ГЕНЕТИКА
ЖИВОТНЫХ
УДК 575.11.113:598.842
НОВЫЙ СЛУЧАЙ РЕКОМБИНАЦИИ МЕЖДУ ЯДЕРНЫМ
И МИТОХОНДРИАЛЬНЫМ ГЕНОМАМИ В РОДЕ Calliope Gould, 1836
(Muscicapidae, Aves): ГИПОТЕЗА ПРОИСХОЖДЕНИЯ
Calliope pectoralis Gould, 1837
© 2019 г. Л. Н. Спиридонова1, *, О. П. Вальчук1, Я. А. Редькин2
1
Федеральный научный центр биоразнообразия Дальневосточного отделения
Российской академии наук, Владивосток, 690022 Россия
2
Научно-исследовательский Зоологический музей Московского государственного университета
им. М.В. Ломоносова, Москва, 125009 Россия
*e-mail: spiridonova@biosoil.ru
Поступила в редакцию 01.02.2018 г.
После доработки 07.03.2018 г.
Принята к публикации 03.04.2018 г.
Впервые предложена гипотеза гибридного происхождения Calliope pectoralis от видов С. calliope и
C. obscura на основе молекулярно-генетических данных и фенотипических признаков. Исследова-
ны 80 образцов С. calliope и один образец С. pectoralis tschebaiewi. Получаемый в результате ПЦР про-
дукт гена цитохрома b, три транспортных РНК, ND6 и контрольный регион (3.2 тпн), у 22 особей
С. calliope имел гетерогенную природу по типу гетероплазмии. Результат клонирования этих ампли-
конов показал два варианта клонов: ген цитохрома b С. calliope и ядерный псевдоген, гомологичный
гену цитохрома b С. pectoralis (сходство 96%). Реконструкция связей мт-гаплотипов гена цитохрома b и
его ядерных копий распределила клонированные последовательности в двух кладах – C. calliope и
C. pectoralis. Мы полагаем, что в данном случае имело место межгеномное рекомбинационное собы-
тие – перенос ядерной копии гена цитохрома b C. calliope в митохонриальный геном у гибридной сам-
ки, ставшей основателем вида С. pectoralis. Второй вид, участвовавший в гибридизации с C. calliope, по
морфологическим признакам, вероятно, был C. obscura, так как только он из всех видов Calliope имеет
черную грудь и черные с белыми основаниями внешние рулевые перья, похожие на таковые у C. pec-
toralis.
Ключевые слова: межвидовая гибридизация, NUMT, мтДНК, межгеномная рекомбинация, Calliope
calliope, C. obscura, C. pectoralis.
DOI: 10.1134/S0016675819010132
Изучение молекулярных маркеров митохон- Ядерные копии мтДНК чаще всего считают
дриальной ДНК с целью видовой идентификации помехами, которые приводят к ошибочным по-
(ДНК-кодирования) привело к появлению целого строениям филогений таксонов разного ранга [4],
ряда “ложных” таксонов и связано в первую оче- что справедливо в тех случаях, когда ядерные копии
редь с ядерными копиями митохондриальных генов амплифицируются у одних таксонов, а митохон-
(NUMT), коамплифицирующимися вместе с орто- дриальные гены у других, поскольку сравниваемые
логичными генами митохондриальной ДНК [1]. последовательности имеют разные источники про-
NUMT считают фрагментами древней мтДНК и исхождения [5–7]. Использование митохондриаль-
предлагают использовать в качестве базальных ных генов и их фрагментов в качестве маркеров
ветвей филогенетических деревьев митохондри- эволюционного процесса некоторыми исследо-
альных маркеров для реконструкции предкового вателями ставится под сомнение, поскольку во
состояния митохондриального генома [2]. Одна- многих случая доказана неравномерность мути-
ко в работе Б.В. Андрианова с соавторами получе- рования митохондриальных генов и их частей [8].
ны данные, указывающие на высокую степень из- Наряду с этим обнаружены факты нарушения ма-
менчивости псевдогенов митохондриального теринского наследования митохондрий, описаны
происхождения после их интеграции в хромосо- рекомбинации мтДНК, гетероплазмия и высокий
мы, что противоречит представлению о них как о уровень индивидуального гаплотипического раз-
“молекулярных ископаемых” [3]. нообразия [9, 10].
81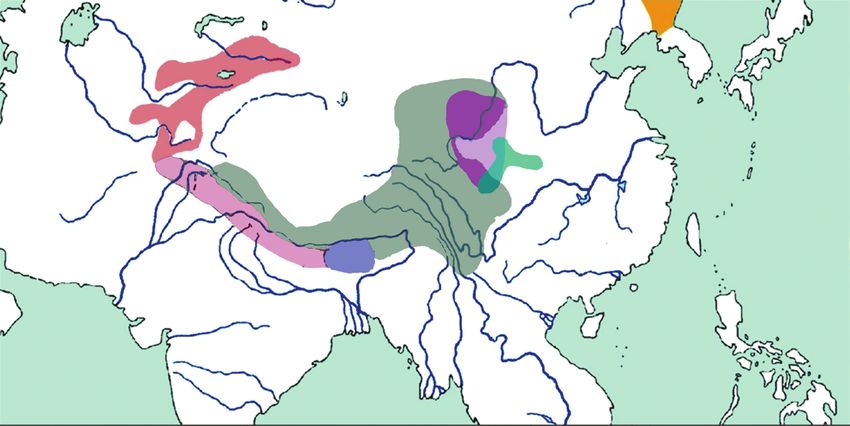
82 СПИРИДОНОВА и др.
1
6
2
7
5 3
4
Рис. 1. Ареалы видов Calliope и их подвидов: C. calliope (1 – C. c. calliope; 2 – С. c. beicki), С. pectoralis (3 – С. p. tschebaiewi;
4 – С. p. confusa; 5 – С. p. pectoralis; 6 – С. p. ballioni), 7 – C. obscura.
Несмотря на общепринятое негативное мне- тае представлен изолированной горной формой
ние о ядерных копиях митохондриальных генов, С. c. beicki (Meise, 1937). Монотипические виды
наши данные показывают в некоторых случаях по- C. pectardens и C. obscura населяют горы Южного и
тенциальную важность использования NUMT для Центрального Китая [15].
уточнения филогенетических построений. Ранее C. pectoralis высокогорный вид, населяющий
при изучении подвидов С. calliope (Pallas, 1776) нами альпийские и субальпийские луга с кустарниковой
была показана роль митохондриальных псевдоге- растительностью, образует ряд хорошо обособлен-
нов как источника новых вариантов подвидовых ных подвидов [14, 16, 18–20]. C. p. pectoralis населя-
гаплотипов мтДНК [11]. Анализ митохондриаль- ет южный макросклон Гималаев от Каракорума до
ных генов и их ядерных копий отдельных предста- восточных районов Непала; C. p. confusa (E. Hartert,
вителей рода Calliope Gould, 1836 выявил новые 1910) обитает восточнее – в северо-восточных частях
факты, подтверждающие важную роль митохон- Индии и Бутане; самая северная форма C. p. ballioni
дриальных псевдогенов в поддержании генетиче- (Severtsov, 1873) занимает горные системы Пами-
ского разнообразия мтДНК. Объектом данного ро-Алая и Тянь-Шаня. Наконец, ареал морфоло-
исследования стал черногрудый соловей-красно- гически обособленной формы C. p. tschebaiewi
шейка C. pectoralis Gould, 1837, проблема проис- Przevalski, 1876, простирается вдоль северного
хождения которого заслуживает специального макросклона Гималаев от южной оконечности
комментария. Каракорума до крайнего севера Мьянмы, Юнна-
Выделение соловьев-красношеек в качестве ни, а также охватывает западную часть провин-
самостоятельного рода Calliope из большой, мор- ции Сычуань, Ганьсу и Восточный Цинхай (см.
фологически очень неоднородной группы “соло- рис. 1).
вьев” Luscinia s. l., основанное на результатах со- Согласно мнению многих исследователей,
временных филогенетических построений [12, таксономический статус C. p. tschebaiewi нуждает-
13], получило широкое признание и было приня- ся в переоценке. Еще Ч. Вори [18] указывал на то,
то в основных современных списках птиц Мира что pectoralis и tschebaiewi могут быть разными ви-
[14, 15]. В состав данного рода вошли политипи- дами. Исследуя вариации размеров и окраски под-
ческие виды C. calliope и C. pectoralis и два моноти- видов С. pectoralis, В.М. Лоскот и К.К. Далецкая
пических вида – C. pectardens David, 1877 и [16] пришли к выводу о наибольшей дивергенции
C. obscura (Berezowski et Bianchi, 1891). Большин- С. p. tschebaiewi от других подвидов, возможно со-
ство представителей рода Calliope населяют гор- ответствующей видовому статусу. С учетом того,
ные районы Азии (рис. 1). что в коллекциях не были найдены особи со строго
Черногрудый красношейка, представленный промежуточными признаками между tschebaiewi и
рядом подвидов, распространен от Тянь-Шаня и другими подвидами, авторы предложили считать
Памиро-Алая, через Гималаи до восточного Цин- эту форму мегаподвидом [16]. Позже П. Расмусс-
хая, Ганьсу и западной части провинции Сычуань эн и Дж. Андертон [20] высказали мнение о про-
[16]. Собственно соловей-красношейка C. calliope, межуточном положении формы C. p. tschebaiewi
широко распространенный на большей части Си- между видами C. calliope и C. pectoralis по морфо-
бири и Дальнего Востока [17], в Центральном Ки- логическим признакам. По мнению этих же авто-
ГЕНЕТИКА том 55 №1 2019НОВЫЙ СЛУЧАЙ РЕКОМБИНАЦИИ 83
ров, поскольку вокализация всех трех таксонов вание клонов на наличие вставки осуществляли с
(calliope, pectoralis и tschebaiewi) очень сходна, в праймерами M13/pUCfw (5'-GCCAGGGTTTTC-
дальнейшем они могут рассматриваться либо в CCAGTCACGA-3') и M13/pUCrv (5'-GAGCGGA-
составе единого политипического вида, либо в TAACAATTTCACACAGG-3') при следующих усло-
качестве трех самостоятельных видов. И наконец, виях: преденатурация – 2 мин/94°C, затем 35 цик-
Я. Лю с соавт. [21] на основании данных мульти- лов: денатурация – 30 с/94°C, отжиг – 30 с/55°С,
локусного анализа по двум митохондриальным и
двум ядерным генам, а также по результатам срав- синтез – 1 мин/72°C, окончательный синтез –
нения вокализации и внешним морфологическим 5 мин/72°C. Все этапы эксперимента были проведе-
отличиям предлагают рассматривать форму tsche- ны дважды, чтобы подтвердить воспроизводимость
baiewi в качестве отдельного монотипического ви- результатов. Клонированные последовательности
да. Данная точка зрения отражена и в последнем гена cytb депонированы в Генбанк ENA/EMBL под
списке воробьинообразных птиц Мира [15]. номерами доступа LT991756–LT991862.
Гипотез о гибридном генезисе черногрудого Секвенирование и компьютерная обработка по-
красношейки С. pectoralis s. l. ранее не высказыва- следовательностей. Продукты амплификации
лось, хотя на явно промежуточный характер при-
были использованы для циклического секвени-
знаков C. p. tschebaiewi некоторые исследователи
обращали специальное внимание [20]. В нашей рования с набором реагентов ABI PRISM®Big-
работе мы рассматриваем гипотезу гибридного DyeTM Terminator v. 3.1. Реакцию присоединения
происхождения С. pectoralis от видов С. calliope и метки проводили с теми же праймерами. Резуль-
С. obscura, основываясь на молекулярных данных и таты прочитывали на автоматическом лазерном
морфологическом облике обcуждаемых таксонов. секвенаторе ABI PRISM 3130 (Applied Biosystems,
Помимо этого представляем очередной случай USA/Hitachi, Japan).
прошедшего рекомбинационного события между
гомологичными последовательностями ядерного Прямые и обратные последовательности были
и митохондриального геномов, в результате кото- собраны в пакете программ Staden 1.53 [22] и вы-
рого возник митохондриальный гаплотип С. pec- ровнены в программе Clustal W, предложенной в
toralis. MEGA ver. 6 [23]. Для анализа нуклеотидного раз-
нообразия клонов и построения матрицы поли-
морфных сайтов использовали программу DnaSP
МАТЕРИАЛЫ И МЕТОДЫ ver. 5.0 [24]. Реконструкция филогенетических
Образцы ДНК и амплификация гена цитохрома b связей между гаплотипами методом максимально
мтДНК. Исследованы 80 образцов С. calliope, от- правдоподобия (Maximum Likelihood, ML) прове-
ловленных в окрестностях станции кольцевания дена также в MEGA.
“Амуро-Уссурийского центра биоразнообразия
птиц” (Приморский край, долина р. Литовка, Секвенирование митохондриального фрагмен-
N 42.962°; E 132.80°) в период осенней миграции та (3.2 тпн) от особи-мигранта № 16 проведено ме-
2011 г., и один образец С. pectoralis tschebaiewi, со- тодом пиросеквенирования на Roche GS Junior.
бранный на севере Мьянмы, штат Качин Приготовление быстрой библиотеки с лигирован-
(N 27.489°; E 97.191°) (сборы В.Н. Сотникова). ными адаптерами, проведение эмульсионной
ДНК выделяли из образцов крови, фиксирован- ПЦР, секвенирование с реагентами GS Junior Ti-
ной в 96%-ном этаноле, с помощью набора QIA- tanium Series выполнялись согласно инструкциям
gen DNeasy® Tissue Kit (Qiagen, Inc.). Мы исполь- фирмы-изготовителя прибора и реактивов (454
зовали последовательности гена cytb из Генбанка Life Science Corp., Roche Company, Branford, CT
NCBI: C. p. pectoralis (Гималаи, KJ456329), C. p. balli-
06405, USA). Работа выполнена на оборудовании
oni (Казахстан, HM633321). Необходимо отме-
тить, что молекулярный анализ мигрантов С. cal- ЦКП “Биотехнология и генетическая инженерия”
liope проводился в отсутствие образцов С. pectoralis. ФНЦ Биоразнообразия ДВО РАН. Нуклеотидные
Амплификация фрагмента митохондриальной последовательности собирали с помощью про-
ДНК (мтДНК), включающего ген cytb, три т-РНК, граммы GS De Novo Assembler (http://454.com/
ND6 и CR, и секвенирование на его основе гена products/analysis-software/index.asp). Секвениро-
cytb проведены, как описано в работе [11]. вание фрагмента прошло со средним покрытием
Клонирование фрагментов. Гетерогенные фраг- ×100–300. Для подтверждения рекомбинационно-
менты восьми особей проклонированы с помо- го события между ядерным и митохондриальным
щью набора InsTAcloneTM PCR cloning Kit (Fermen- геномами проведено тестирование ядерного псев-
tas, Litva) согласно инструкции производителя. догена и соответствующих фрагментов мтДНК
Для легирования применили вектор pTZ57R/T. C. calliope, C. pectoralis, Luscinia svecica (Linnaeus,
Трансформацию проводили в компетентные 1758), Larvivora akahige (Temminck, 1835) и Tarsiger
клетки Escherichia coli штамма XL1-Blue. Тестиро- cyanurus (Pallas, 1773) в программе RDP ver. 4 [25].
ГЕНЕТИКА том 55 №1 201984 СПИРИДОНОВА и др.
13 600 13 800 14 K 14 200 14 400 14 600 14 800 15 K 15 200 15 400 15 600 15 800 16 K 16 200 16 400 16 600
Genes
tRNA-Thr ND6
ADU57283.1
CYTB tRNA-Pro tRNA-Glu
ADU572…
misc_feature Features
Control region
Рис. 2. Схема амплифицированного фрагмента митохондриальной ДНК, включающего ген cytb, т-РНК, ND6 и кон-
трольный регион (D-петля) (3.2 тпн), из которого был секвенирован ген cytb (1141 пн).
РЕЗУЛЬТАТЫ ДНК. Картина распределения нуклеотидов в ядер-
Причиной появления данного исследования ных псевдогенах calliope и гене cytb С. pectoralis име-
стали рутинные эксперименты подбора условий ет хорошо выраженную гомологию, что указыва-
амплификации длинных фрагментов для даль- ет на их общее происхождение. В то же время не-
нейшего использования на секвенаторе нового обходимо отметить и значительную дивергенцию
поколения Roche 454. Анализ фрагмента длиной между ядерным псевдогеном C. calliope и митохон-
около 3.2 тнп, включающего ген cytb, три т-РНК, дриальным геном cytb С. pectoralis, что обусловлено
ND6 и CR (рис. 2), выявил в последовательностях длительным пребыванием паралога в ядерном ге-
22 из 80 исследуемых особей С. calliope присут- номе после рекомбинационного события.
ствие двойных пиков по типу гетероплазмии. В На рис. 4 показаны отношения видов C. calliope
результате клонирования ампликонов от восьми и C. pectoralis и клонированных последовательно-
особей получены 162 клона гена cytb, в среднем по стей C. calliope. Реконструкция связей мт-гаплоти-
20 клонов от каждого фрагмента. пов гена cytb и его ядерных копий методом макси-
Последовательности клонов сравнивали с ра- мального правдоподобия показала, что все клоны
нее выявленными гаплотипами подвидов С. calli- распределились в трех кладах – C. calliope (I и II ва-
ope и с последовательностями, хранящимися в базе риант) и C. pectoralis (III вариант). Полученный
данных Генбанка NCBI. Картина распределения результат указывает на родственную связь ядер-
гетероплазмичных сайтов в данном случае оказа- ного псевдогена С. calliope и митохондриального
лась иной, чем в предыдущих анализах [11]. Один гена С. pectoralis. Поскольку ядерный паралог и
вариант клонов у всех птиц оказался геном cytb митохондриальный ген находятся в разных гено-
мтДНК С. calliope, а второй по распределению мах, то мы полагаем, что имело место межгеном-
фиксированных мутаций был сходным на 96% с ное рекомбинационное событие, как и в случае
геном cytb С. pectoralis. происхождения подвидовых гаплотипов C. calli-
ope [11, 17].
В некоторых клонированных последователь-
ностях обнаружены митохондрильные стоп-ко- Для подтверждения рекомбинационного со-
доны (TAA), единичные делеции, приводящие к бытия между ядерным псевдогеном и мтДНК бы-
сдвигу рамки считывания, и 11 терминирующих ло проведено тестирование NUMT и фрагментов
TGA-кодонов (“opal” или “umber”). Анализ клони- мтДНК C. calliope, C. pectoralis, Luscinia svecica,
рованных последовательностей выявил высокий Larvivora akahige и Tarsiger cyanurus в программе
уровень изменчивости, обусловленный хаотично RDP ver. 4. В результате фрагмент cytb–CR
распределенными единичными мутациями и мно- мтДНК C. pectoralis оказался рекомбинантным с
гочисленными рекомбинационными перестрой- достоверными значениями вероятности (Chimae-
ками между разными вариантами гаплотипов, ra, P = 1.458 × 10–04; MaxChi, P = 3.322 × 10–05;
представленных в ядерном геноме С. calliope (рис. 3). GENECONV, P = 1.123 × 10–08; SiScan, P = 5.562 ×
Например, длина рекомбинантных участков в × 10–13).
клонированных фрагментах варьировала от 85 пн
у клона 16.6 (образец № 16) до 869 пн у клона 6.2
(образец № 6). Необходимо особо отметить, что ОБСУЖДЕНИЕ
вставки других вариантов гаплогрупп, обнару- Популяционные исследования митохондри-
женные в ядерных копиях митохондриальных ге- альных маркеров у многих таксонов птиц выявля-
нов у одной особи, свидетельствуют о высокой ют две филогеографические картины. Одна из них
скорости изменчивости псевдогенов благодаря ре- характеризуется отсутствием хорошо выраженной
комбинационным процессам самой ядерной филогеографической структуры и описана, на-
ГЕНЕТИКА том 55 №1 2019matrix 10 20 30 40 50 60 70 80 90 100 110 120 130 140 150 160 170 180
ГЕНЕТИКА
LK932558 * * * * * * * * * * * * * * * * * *
LK932560
C. calliope LK932557
LK932690
LK932656
KJ456329
том 55
C. pectoralis HM633321
tschebaiewi
'6.8'
'6.10'
'16.1'
'16.2'
№1
'25.7'
'25.1'
'25.3'
'25.4'
'25.5'
'25.6'
'25.11'
2019
Клоны '25.12'
'25.13'
C. calliope, '25.14'
'29.1'
вариант III '29.2'
'45.10'
'45.8'
'45.9'
'45.12'
'45.13'
'45.18'
'45.19'
'47.16'
'47.8'
'47.10'
'47.11'
'47.14'
'47.15'
'47.18'
'47.17'
'47.12'
'47.13'
'6.13'
'6.5'
'45.15'
'16.8'
Клоны '16.5'
'16.4'
С. calliope, '6.7'
'6.2'
вариант I '16.3'
'16.6'
'16.7'
'25.8'
НОВЫЙ СЛУЧАЙ РЕКОМБИНАЦИИ
'6.11'
'6.12'
'6.15'
'6.4'
'6.1'
Клоны '45.14'
'45.11'
C. calliope, '29.3'
'6.6'
вариант II '6.9'
'21.4'
'21.5'
'21.6'
'21.7'
Рис. 3. Матрица полиморфных сайтов гена cytb мтДНК C. calliope и C. pectoralis и клонированных ядерных копий C. calliope. Серым цветом выделены вертикально
сайты, идентичные для C. pectoralis и ядерных копий C. calliope, горизонтально – рекомбинантные участки вариантов ядерного псевдогена C. calliope. Цифрами
обозначены номера птиц-мигрантов C. calliope и их клонов.
8586
matrix 190 200 210 220 230 240 250 260 270 280 290 300 310 320 330 340 350 360 370 380
LK932558
* * * * * * * * * * * * * * * * * * * *
LK932560
C. calliope LK932557
LK932690
LK932656
KJ456329
C. pectoralis HM633321
tschebaiewi
'6.8'
'6.10'
'16.1'
'16.2'
'25.7'
'25.1'
'25.3'
'25.4'
'25.5'
'25.6'
'25.11'
'25.12'
Клоны '25.13'
'25.14'
C. calliope, '29.1'
'29.2'
вариант III '45.10'
'45.8'
'45.9'
'45.12'
'45.13'
'45.18'
'45.19'
'47.16'
'47.8'
'47.10'
'47.11'
'47.14'
'47.15'
'47.18'
'47.17'
'47.12'
'47.13'
'6.13'
'6.5'
'45.15'
'16.8'
СПИРИДОНОВА и др.
Клоны '16.5'
'16.4'
С. calliope, '6.7'
'6.2'
вариант I '16.3'
'16.6'
'16.7'
'25.8'
'6.11'
'6.12'
'6.15'
'6.4'
Клоны '6.1'
'45.14'
C. calliope, '45.11'
'29.3'
вариант II '6.6'
'6.9'
ГЕНЕТИКА
'21.4'
'21.5'
'21.6'
'21.7'
Рис. 3. Окончание.
том 55
№1
2019НОВЫЙ СЛУЧАЙ РЕКОМБИНАЦИИ 87
C. c. anadyrensis
C. c. camtschatkensis
Клоны гена cytb – NUMT (I)
C. calliope
C. c. sachalinensis
100
C. c. calliope –
Клоны гена cytb – NUMT (II)
16.6
100 HM633321 C. p. ballioni
99 KJ456329 C. p. pectoralis
C. p. tschebaewi
C. pectoralis
Клоны NUMT (III) C. calliope
100
Рис. 4. Реконструкция филогенетических связей митохондриальных гаплотипов С. pectoralis и ядерных копий мито-
хондриальных генов С. calliope, построенная по методу ML. I, II и III – варианты клонов.
пример, для Parus major s.l., P. montanus s.l. [26], довая гибридизация, в результате которой появля-
Perisoreus infaustus, Nucifraga caryocatactes [27], Nu- ется новый вариант гаплотипа, не свойственный
cifraga columbiana [28], Luscinia svecica [29]. Другая данному таксону. Однако сходство распределения
картина, наоборот, имеет хорошо выраженную полиморфных сайтов между мтДНК и ее ядерны-
филогеографическую структуру и обнаружена у ми копиями невозможно объяснить ни одним из
C. calliope [17], Motacilla flava s.l., M. citreola s.l. [30], этих путей. Третий путь, который здесь рассматри-
Ficedula parva s.l. и Alauda arvensis s.l. [31], Pinicola вается, – обычная гомологичная рекомбинация
enucleator [32]. Причины таких явлений обычно между ядерным и митохондриальным геномами,
объясняются событиями ледникового периода. которая внезапно приводит к смене мт-гаплотипа
Комплекс C. pectoralis по данным митохондриаль- на новый, уже имеющийся в ядерном геноме пара-
ного маркера (cytb) характеризуется хорошо вы- лог мтДНК.
раженной филогеографической структурой и, по
мнению Я. Лю с соавторами, отделился от бли- Кроссинговер между гомологичными участками
жайшего родственника, C. calliope, распростра- ядерного и митохондриального геномов и перенос
ненного севернее, в период раннего плиоцена ядерной копии гена cytb в мтДНК мог произойти в
(3.2–4.8 млн лет назад) [21]. геноме гибридной самки, ставшей основателем ви-
да С. pectoralis. Следовательно, C. pectoralis получил
Ранее на примере соловья-красношейки мы свой мт-гаплотип из ядерных псевдогенов C. cal-
показали альтернативные пути формирования liope (одного из родителей). Транспорт нуклеино-
филогеографической структуры вида с участием вых кислот различной длины в митохондрии и
ядерных псевдогенов [17]. Наши данные предпо- механизмы, с помощью которых происходят эти
лагают аналогичный сценарий формирования процессы, описаны в литературе [33, 34]. В обна-
филогеографической структуры и для комплекса руженном рекомбинационном событии нам пока
C. pectoralis. Обнаруженные у C. calliope ядерные не удалось определить полную длину рекомби-
копии гена cytb мтДНК сходны с митохондриаль- нантного фрагмента, которая может оказаться
ным геном С. pectoralis. Как можно объяснить та- весьма значительной, как было установлено на
кую гомологию последовательностей из разных примере домашней кошки [35]. Однако присут-
геномов и у разных таксонов? Теоретически су- ствие в нем гена cytb и контрольного региона (CR)
ществуют два пути формирования дивергирован- установлено экспериментально.
ных гаплотипов мтДНК: первый – постепенное
накопление замен в результате длительного пре- Рекомбинационное событие, возможно, про-
бывания популяций в изоляции и второй – межви- изошло во время или вскоре после межвидовой
ГЕНЕТИКА том 55 №1 201988 СПИРИДОНОВА и др.
SYSb0772 Yunnan
SYSb1085 Tibet
Calliope pectoralis tschebaiewi
SYSb1086 Tibet
–/99
SYSb0771 Yunnan
SYSb1528 Hebei
93/100 SYSb2545 Gansu
–/96
DZUG-U660 Japan
C. calliope
SYSb1527 Hebei
72/100 SYSb1524 Hebei
SYSb1525 Hebei
SYSb1090 Tajikistan
SYSb0024 Xinjiang
82/100 SYSb1091 Tajikistan
SYSb1089 Tajikistan C. p. ballioni
DZUG-U30 Kazakhstan
–/100
SYSb1088 Tajikistan
SYSb0462 Kazakhstan
SYSb0773 Yunnan
SYSb0770 Yunnan
C. p. tschebaiewi
SYSb0540 Qinghai
SYSb0774 Yunnan
98/100 Shaanxi C. obscura
SYSb0463 Sichuan
C. pectardens
DZUG-U40 Sichuan 0.005
Tarsiger johnstoniae
Рис. 5. Филогенетические взаимосвязи четырех видов Calliope, построенные на основе объединенных последователь-
ностей ядерного миоглобина и ODC гена (1364 пн) [21].
гибридизации на периферии современного ареа- данными и подтверждает гипотезу гибридного
ла C. calliope beicki и ареала С. obscura. Низкая чис- происхождения вида С. pectoralis от C. calliope.
ленность особей, вероятнее всего в районе, где Межвидовая гибридизация у птиц случается
располагается их современная зона контакта c достаточно часто и отмечена примерно для 850
подвидом C. p. tschebaiewi, способствовала после- видов [36]. Однако обособление межвидовых ги-
дующему распространению рекомбинантного бридов в отдельный вид и возникновение преко-
мт-гаплотипа по типу эффекта основателя (см. пуляционных изолирующих механизмов случает-
рис. 1). Это дало возможность новому гаплотипу ся редко. Не так давно в результате естественной
закрепиться и стать таксон-специфичным по ме- гибридизации большого кактусового земляного
ре заселения новых территорий. Обратное пред- вьюрка Geospiza conirostris со средним земляным
положение о внедрении митохондриального гап- вьюрком G. fortis в течение нескольких поколений
лотипа C. p. tschebaiewi в ядерный геном C. calliope сформировался новый вид дарвиновых вьюрков
представляется маловероятным по нескольким [37]. Особи нового вида птиц отличаются от роди-
причинам. Во-первых, из-за значительной уда- тельских видов формой и размерами клюва, име-
ленности мест, где был обнаружен ядерный псев- ют другую вокализацию и спариваются только с
доген (российский ДВ), и, во-вторых, высоким представителями своего вида. Вероятно таким же
гнездовым консерватизмом этих видов соловьев. путем произошел комплекс С. pectoralis.
На рис. 5 показаны отношения видов Calliope По данным митохондриальных маркеров вре-
по двум ядерным генам, взятые из работы Лю с мя дивергенции по “молекулярным часам” между
соавт. [21]. Особенно хочется обратить внимание гаплотипами С. calliope и С. pectoralis оценено в
на разделение комплекса С. pectoralis на две филе- 4 млн лет [21]. Результаты нашего исследования
тические линии, одна из которых близка к С. cal- ставят под сомнение возможность использования
liope, а вторая незначительно дивергировала, од- “молекулярных часов”, поскольку происходит не
нако обе находятся в кладе последнего вида. Дан- только резкая смена гаплотипа, но накопление
ный результат полностью согласуется с нашими мутаций идет вначале в ядерном геноме. Сравни-
ГЕНЕТИКА том 55 №1 2019НОВЫЙ СЛУЧАЙ РЕКОМБИНАЦИИ 89
C. p. pectoralis
× (II)
C. obscura
× C. p. tschebaiewi
(I)
C. calliope
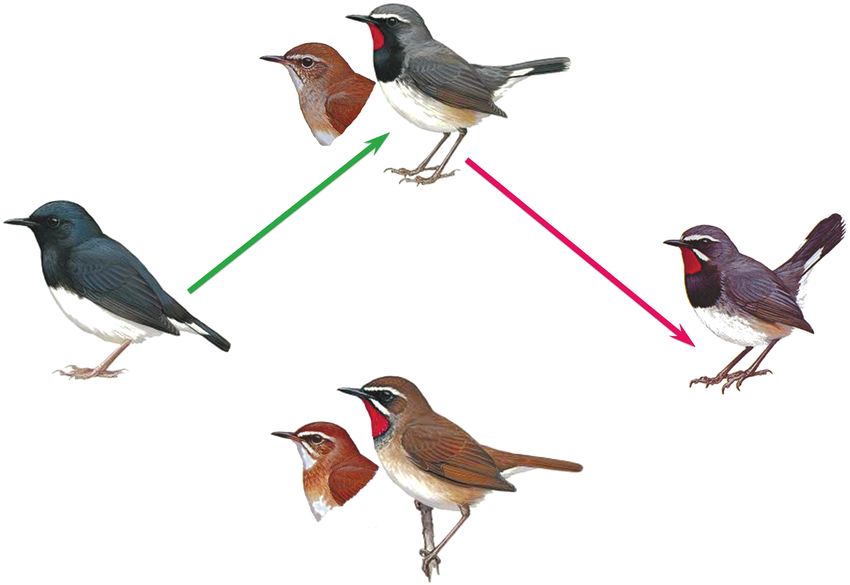
Рис. 6. Схема двух событий межвидовой гибридизации (I и II) с полом птиц каждого родительского вида, в результате
которых возникли подвиды C. pectoralis. Зеленая стрелка указывает форму, возникшую в результате гибридизации (I);
красная стрелка – форму, возникшую в результате гибридизации (II). (Цветные фотографии см. статью в электронном
виде.)
вая генетическое сходство (96%) ядерных копий этом у взрослых особей окраска пигментирован-
мтДНК С. calliope и митохондриального гаплоти- ных частей рулевых перьев полностью черная.
па cytb С. pectoralis со случаем, описанным в на- Для самцов черногорлого соловья свойственна
шей предыдущей работе [11], можно лишь отме- полностью черная окраска боков головы, горла и
тить, что становление последнего вида в резуль- груди, образующая широкий черный пластрон,
тате гибридизации произошло раньше, чем резко контрастирующий с преимущественно бе-
возникновение подвидов С. calliope, поскольку лой окраской живота.
сходство гена cytb и его ядерных копий у С. calliope
выше (до 100%) по сравнению с таковыми у С. pec- Соловей-красношейка C. calliope отличается
toralis. наличием у самцов резко выраженного цветного
пластрона (образованного перьями с ярко-крас-
Помимо генетических данных, указывающих ными вершинами и белыми основаниями), охва-
на гибридное происхождение С. pectoralis, мы
тывающего подбородок, горло и верхнюю часть
провели фенотипическое сравнение возможных
зоба, резко оконтуренного черными перьями че-
участников гибридизации, в результате которой
возник этот вид. По внешним морфологическим люстного штриха по бокам горла, переходящего в
признакам представители рода Calliope отличают- серый оттенок на боках зоба и груди. Для обоих
ся достаточно четко, в первую очередь благодаря полов характерна своеобразная топография
особенностям окраски оперения (рис. 6). Для окраски головы, обусловленная наличием четко
черногорлого соловья C. obscura характерна чер- выраженных светлых бровей (над глазом и крою-
новато-синяя окраска верха у взрослых самцов, щими уха), а также светлых “усов” (продольных
отсутствие депигментированных участков по бо- полос, протягивающихся от основания подклю-
кам темени (“белых бровей”) и по бокам горла вья ниже темного оперения уздечки и уха). Как
(“усов”), наличие широких белых полей в осно- для самцов, так и для самок характерна оливково-
вании рулевых перьев (за исключением централь- бурая окраска верхней стороны головы и тела, од-
ной пары) и относительно короткий хвост. При ноцветная (без белых полей) окраска рулевых то-
ГЕНЕТИКА том 55 №1 201990 СПИРИДОНОВА и др.
го же оттенка, а также относительно большая Светлая бровь над глазом в среднем длиннее, чем
длина хвоста. у черногрудого красношейки. Ниже кроющих
уха, по бокам горла хорошо выражена белая поло-
Для подвидов черногрудого красношейки
са (ус), совершенно сходная с таковой у C. calliope.
С. pectoralis свойственны внешние признаки, в
Окраска верхней стороны тела серовато-оливко-
различной степени переходные между черногор-
вая у самцов и оливково-бурая у самок, в наи-
лым соловьем и соловьем-красношейкой. Подвиды
большей степени приближающаяся к расцветке
С. p. pectoralis, С. p. ballioni и C. p. confusa обладают
верхней стороны тела соловья-красношейки. Бе-
оливково-голубовато-серой окраской верхней
лые пятна на вершинах рулевых мельче, чем у
стороны тела; у самцов на подбородке и горле вы-
других подвидов С. pectoralis, или отсутствуют у
ражено относительно небольшое красное пятно,
самок и молодых птиц [16]. Хвост относительно
со всех сторон окруженное полностью черным
длиннее, чем у других черногрудых красношеек, а
пластроном (перья уздечки, передней и нижней
в целом по своим пропорциям C. p. tschebaiewi
частей кроющих уха, боков горла, зоба и большей
практически сходен с китайским подвидом C. cal-
части груди), у обоих полов выражена белая
liope beicki. Таким образом, форма tschebaiewi объ-
бровь, которая при этом заметно короче, чем у
ективно имеет большее морфологическое и фе-
C. calliope, и заканчивается сразу позади глаза.
нотипическое сходство с C. calliope, чем другие
Хвост длиннее, чем у C. obscura, но в среднем ко-
подвиды С. pectoralis, что косвенно указывает на
роче, чем у соловья-красношейки. У самцов руле-
ее гибридное происхождение.
вые центральной пары целиком буровато-серые,
а пигментированные части прочих перьев хвоста По нашему мнению, разделение pectoralis на
черные, тогда как основания большинства из них две филогенетические линии можно объяснить
белые, при этом ширина депигментированных двумя фактами гибридизации, схема которых
участков в среднем меньше, чем у черногорлого представлена на рис. 6. На первом этапе произо-
соловья. Уникальной чертой окраски всех черно- шла гибридизация самки С. calliope, несущей ре-
грудых красношеек является наличие округлых комбинантный гаплотип мтДНК, с самцом C. ob-
белых пятен на вершинах большинства рулевых scura (см. рис. 6). Результатом этого случая стало
перьев (за исключением центральной пары), в возникновение формы, предковой для pectoralis,
меньшей степени выраженные или отсутствую- ballioni и confusa, которые имеют более широкий
щие у самок и молодых птиц [16]. черный пластрон на груди, сходный с таковым и
видимо унаследованный от C. obscura, а также
Вторым родительским видом С. pectoralis, судя красное пятно на горле и белые брови, приобре-
по морфологическим признакам, мог быть C. ob- тенные от С. calliope. На втором этапе участника-
scura. Из всех видов соловьев только этот вид име- ми гибридизации были самец С. calliope и самка
ет черные горло и грудь. C. pectoralis сохранил фе- С. pectoralis. Потомство этой пары несло уже
нотипические признаки обоих предполагаемых двойную дозу ядерного генома С. calliope, что от-
родителей. Самцы имеют красное горло и белые ражено на дереве, построенном по ядерным ге-
брови (C. p. tschebaiewi еще и белые усы) от C. calliope; нам (см. рис. 5) [21]. Результатом этого случая
черную грудь и черные, с белыми основаниями, стало возникновение формы tschebaiewi, у кото-
внешние рулевые перья от C. obscura. Самки похожи рой морфологические признаки С. calliope про-
на самок C. calliope, но более темные и серые. Моло- явились значительно сильнее, чем у прочих форм
дые птицы похожи на таковых C. calliope. Отличает- черногрудой красношейки.
ся C. pectoralis как от C. calliope, так и от C. obscura бе-
Таким образом, мы обнаружили новые факты,
лыми кончиками рулевых, характерными только
указывающие на гомологичную рекомбинацию
для представителей комплекса черногрудых крас-
между участками митохондриального и ядерного
ношеек.
геномов у соловья-красношейки, в результате ко-
C. p. tschebaiewi морфологически резко отлича- торой на основе последовательностей ядерных
ется от других подвидов черногрудой красношей- псевдогенов возникли новые митохондриальные
ки, приближаясь по целому ряду признаков к гаплотипы гибридного вида C. pectoralis. По моле-
C. calliope. Красное пятно у самцов этой формы кулярным данным и фенотипическим признакам
заметно больше, чем у всех других подвидов чер- C. pectoralis является межвидовым гибридом
ногрудого красношейки (помимо подбородка и C. calliope и C. obscura, а его проанализированный
горла охватывает верхнюю часть зоба), и сходно митохондриальный гаплотип имеет ядерное про-
по размерам с таковым у C. calliope. Охватываю- исхождение. Это новый случай межгеномной ре-
щий цветовое пятно черный пластрон на груди комбинации, установленный нами для рода Cal-
значительно меньше, чем у других С. pectoralis. liope. Обнаруженные варианты ядерных копий
ГЕНЕТИКА том 55 №1 2019НОВЫЙ СЛУЧАЙ РЕКОМБИНАЦИИ 91
гена cytb С. calliope (см. рис. 3) являются потенци- СПИСОК ЛИТЕРАТУРЫ
альным источником новых гаплотипов мтДНК и 1. Song H., Buhay J.E., Whiting M.F., Crandall K.A. Many
открывают скрытый грандиозный запас молеку- species in one: DNA barcoding overestimates the num-
лярной изменчивости мтДНК, которая может ber of species when nuclear mitochondrial pseudogenes
быть реализована через межгеномную рекомби- are coamplified // Proc. Natl Acad. Sci. USA. 2008.
нацию. V. 105. P. 13486–13491. doi 10.1073/pnas.0803076105
Наши данные показывают потенциальную 2. Bensasson D., Zhang D.-X., Hartl D.L., Hewitt G.M. Mi-
важность использования NUMT для правильного tochondrial pseudogenes: Evolution’s misplaced witness-
es // TRENDS in Ecol. Evol. 2001. V. 16. P. 314–321.
анализа филогенетических построений. Изуче-
ние ядерных паралогов в дальнейшем позволит 3. Андрианов Б.В., Романов Д.А., Горелова Т.В. и др.
переосмыслить их роль в формировании генети- Перенос митохондриальной ДНК в ядерный геном
ческого разнообразия в природе и оценке скоро- клеток пересеваемой клеточной линии Drosophila vir-
ilis // Генетика. 2013. Т. 49. № 6. С. 788–792.
сти микроэволюционных процессов по митохон-
дриальным маркерам. Новый факт переноса 4. Bernt M., Braband A., Schierwater B., Stadler P.F. Ge-
ядерных копий мтДНК в митохондриальный ге- netic aspects of mitochondrial genome evolution //
ном посредством симметричной рекомбинации Mol. Phyl. Evol. 2013. V. 69. P. 328–338. doi 10.1016/
j.ympev.2012.10.020
является подтверждением взаимодействия ядер-
ного и митохондриального геномов как единой 5. Arctander P. Comparison of a mitochondrial gene and a
сопряженной генетической системы клетки. corresponding nuclear pseudogene // Proc. R. Soc.
Lond. 1995. V. B262. P. 13–19.
В данной работе впервые подтверждено моле- 6. Zhang D.-X., Hewitt G.M. Nuclear integrations: chal-
кулярными и морфологическими данными ги- lenges for mitochondrial DNA markers // Trends Evol.
бридное происхождение С. pectoralis с механиз- Ecol. 1996. V. 11. P. 247–251.
мом симметричной рекомбинации фрагментами 7. Triant D.A., DeWoody J.A. The occurrence, detection,
ДНК между ядром и митохондриями, а также по- and avoidance of mitochondrial DNA translocations in
казана роль NUMT – ядерных копий мтДНК – в mammalian systematics and phylogeography //
качестве источников новых таксон-специфич- J. Mamm. 2007. V. 88. P. 908–920.
ных митохондриальных гаплотипов. 8. Гречко В.В. Проблемы молекулярной филогенети-
Авторы выражают благодарность А.П. Крюко- ки на примере отряда чешуйчатых рептилий (отряд
ву и М.М. Козыренко за ценные замечания и по- Squamata): митохондриальные ДНК-маркеры //
мощь в редактировании рукописи и С.В. Шедько Мол. биол. 2013. Т. 47. № 1. С. 61–82.
за сборку последовательности NUMT. Авторы 9. Grzybowski T., Malyarchuk B.A., Czarny J. et al. High
выражают глубокую признательность В.М. Лос- level of mitochondrial DNA heteroplasmy in single hair
коту за предоставленную возможность ознаком- roots: reanalysis and revision // Electrophoresis. 2003.
ления с коллекционными материалами Зоологи- V. 24. P. 1159–1165.
ческого института РАН (г. Санкт-Петербург), а 10. Kraytsberg Y., Schwartz M., Brown T.A. et al. Recombi-
также В.Н. Сотникову, собравшему и предоста- nation of human mitochondrial DNA // Science. 2004.
вившему для исследования экземпляр C. p. tsche- V. 304. P. 981.
baiewi из Мьянмы. За помощь в подготовке карты 11. Спиридонова Л.Н., Редькин Я.А., Вальчук О.П., Крю-
гнездовых ареалов соловьев мы чрезвычайно ков А.П. Ядерные копии митохондриальных генов –
признательны А.А. Мосалову. источник новых вариантов гаплотипов гена цито-
хрома b Luscinia calliope (Muscicapidae, Aves) // Ге-
Работа выполнена при поддержке Российско- нетика. 2016. Т. 52. № 9. С. 1069–1080. doi 10.7868/
го фонда фундаментальных исследований (грант S0016675816090137
№ 16-04-01304). Значительная часть использован- 12. Sangster G., Alstrom P., Forsmark E., Olsson U. Multilo-
ных материалов была подготовлена и депонирова- cus phylogenetic analysis of Old World chats and fly-
на в Зоологический музей МГУ при поддержке catchers reveals extensive paraphyly at family, subfami-
гранта Российского научного фонда “Научные ly and genus level (Aves: Muscicapidae) // Mol. Phylo-
основы создания национального банка-депози- genet. Evol. 2010. V. 57. P. 380–392. doi 10.1016/
тария живых систем” (грант № 14-50-00029). j.ympev.2010.07.008
Часть работы, выполненная Я.А. Редькиным, 13. Alstrom P., Song G., Zhang R. et al. Taxonomic status of
осуществлена в рамках государственной темы Blackthroat Calliope obscura and Firethroat C. pec-
AAAA-A16-116021660077-3 – “Таксономический tardens // Forktail. 2013. V. 29. P. 94–99.
и биохорологический анализ животного мира как 14. Dickinson E.C., Christidis L. The Howard & Moore
основа изучения и сохранения структуры биоло- Complete Checklist of the Birds of the World: Passer-
гического разнообразия”. ines. 4th ed., V. 2. Eastbourne: Aves Press, 2014. 752 p.
ГЕНЕТИКА том 55 №1 201992 СПИРИДОНОВА и др.
15. del Hoyo J., Collar N.J. HBW and BirdLife Internation- 27. Haring E., Gamauf A., Kryukov A. Phylogeographic
al Illustrated Checklist of the Birds of the World. V. 2. patterns in widespread corvid birds // Mol. Phyl. Evol.
Passerines. Barcelona: Lynx Edicions, 2016. 904 p. 2007. V. 45. P. 840–862.
16. Loskot V.M., Daletskaya K.K. Plumages and size varia- 28. Dohms K.M., Burg T.M. Molecular markers reveal lim-
tion of the Himalayan rubythroat, Luscinia pectoralis ited population genetic structure in a North American
(Gould, 1837) (Aves: Muscicapidae) // Zoosystematica corvid, Clark’s nutcracker (Nucifraga columbiana) //
Rossica. 2001. V. 9. P. 463–486. PLoS One. 2013. V. 8. P. e79621.
17. Спиридонова Л.Н., Вальчук О.П., Редькин Я.А. и др. 29. Zink R.M., Drovetski S.V., Questiau S. et al. Recent evo-
Филогеография и демографическая история соло- lutionary history of the bluethroat (Luscinia svecica)
вья-красношейки Luscinia calliope // Генетика. across Eurasia // Mol. Ecol. 2003. V. 12. P. 3069–3075.
2017. Т. 53. № 8. С. 933–951. doi 10.7868/
S0016675817080100 30. Pavlova A., Zink R.M., Drovetski S.V. et al. Phylogeo-
graphic patterns in Motacilla flava and M. citreola: spe-
18. Vaurie Ch. The Birds of the Palearctic Fauna. Order cies limits and population history // Auk. 2003. V. 120.
Passeriformes. L.: Witherby, 1959. 762 p. P. 744–758.
19. Collar N. Family Turdidae. Handbook of the Birds of
31. Zink R.M., Pavlova A., Drovetski S., Rohwer S. Mito-
the World, Cuckoo-Shrikes to Thrushes. V. 10 / Eds
chondrial phylogeographies of five widespread Eurasian
Del Hoyo J., Elliott A., Christie D. Barcelona: Lynx
bird species // J. Ornithol. 2008. V. 149. P. 399–413.
Edicions, 2005. P. 514–807.
20. Rasmussen P.C., Anderton J.C. Birds of South Asia. The 32. Drovetski S.V., Zink R.M., Ericson P.G.P., Fadeev I.V. A
Ripley Guide. 2. Attributes and Status. 2nd ed. The multilocus study of pine grosbeak phylogeography sup-
Smithsonian Institution, Michigan State University ports the pattern of greater intercontinental divergence
and Lynx Edicions, Washington D.C., Michigan and in Holarctic boreal forest birds than in birds inhabiting
Barcelona, 2012. 684 p. other high-latitude habitats // J. Biogeog. 2010. V. 37.
P. 696–706.
21. Liu Y., Chen G., Huang Q. et al. Species delimitation of
the white-tailed rubythroat Calliope pectoralis complex 33. Weber-Lotfi F., Koulintchenko M.V., Ibrahim N. et al.
(Aves, Turdidae) using an integrative taxonomic ap- Nucleic acid import into mitochondria: New insights
proach // J. Avian Biol. 2016. V. 47. P. 001–012. into the translocation pathways // Biochim. Biophys.
Acta. 2015. V. 1853. P. 3165–3181. doi 10.1016/j.bbam-
22. Bonfield J.K., Smith K.F., Staden R. A new DNA se-
cr.2015.09.011
quence assembly program // Nucl. Acids Res. 1995.
V. 23. P. 4992–4999. 34. Константинов Ю.М., Дитриш А., Вебер-Лотфи Ф.
23. Tamura K., Stecher G., Peterson D. et al. MEGA6: Mo- и др. Импорт ДНК в митохондрии // Биохимия.
lecular Evolutionary Genetics Analysis version 6.0 // 2016. Т. 81. № 10. С. 1307–1321.
Mol. Biol. Evol. 2013. V. 30. P. 2725–2729. 35. Lopez J.V., Yuhki N., Masuda R. et al. Numt, a recent
24. Librado P., Rozas J. DnaSP v5: A software for compre- transfer and tandem amplification of mitochondrial
hensive analysis of DNA polymorphism data // Bioin- DNA to the nuclear genome of the domestic cat //
formatics. 2009. V. 25. P. 1451–1452. J. Mol. Evol. 1994. V. 39. P. 174–190.
25. Martin D.P., Williamson C., Posada D. RDP2: recombi- 36. Randler C. Avian hybridization, mixed pairing and fe-
nation detection and analysis from sequence align- male choice // Animal Behavi. 2002. V. 63. № 1.
ments // Bioinformatics. 2005. V. 21. № 2. P. 260–262. P. 103–119.
26. Pavlova A., Rohwer S., Drovetski S.V., Zink R.M. Differ- 37. Lamichhaney S., Han F., Webster M.T. et al. Rapid hy-
ent post-Pleistocene histories of Eurasian parids // brid speciation in Darwin’s finches // Science. 2017.
J. Heredity. 2006. V. 97. P. 389–402. doi 10.1126/science.aao4593
A New Case of Recombination between Nuclear and Mitochondrial Genomes
in the Genus Calliope Gould, 1836 (Muscicapidae, Aves):
The Hypothesis of Origin Calliope pectoralis Gould, 1837
L. N. Spiridonovaa, *, O. P. Valchuka, and Ya. A. Red’kinb
a
Federal Scientific Center of the East Asia Terrestrial Biodiversity,
Far East Branch, Russian Academy of Sciences, Vladivostok, 690022 Russia
bZoological Museum, Lomonosov Moscow State University, Moscow, 125009 Russia
*e-mail: spiridonova@biosoil.ru
For the first time we propose a hypothesis of hybrid origin of Calliope pectoralis from two species – C. calliope
and C. obscura based on molecular genetic data and phenotypic characters. We examined 80 samples of
ГЕНЕТИКА том 55 №1 2019НОВЫЙ СЛУЧАЙ РЕКОМБИНАЦИИ 93 C. calliope and one sample of С. pectoralis tschebaiewi. The resulting products of cytochrome b gene, as well as three transport RNAs, ND6 and a control region (3.2 kb) were heterogeneous in 22 specimens of C. calli- ope. The result of cloning of these amplicons showed two variants of clones: the cytochrome b gene of C. cal- liope and the nuclear pseudogen homologous to the cytochrome b gene of C. pectoralis (96% match). Cloned sequences of the cytochrome b gene of mtDNA and its nuclear copies were distributed by reconstruction of connections into two clades – C. calliope and C. pectoralis. We believe that in this case there was an interge- nomic recombination event – namely, the transfer of C. calliope’s nuclear copy of the cytochrome b gene into a mitochondrial genome of a hybrid female that later became the founder of C. pectoralis species. According to morphological features, the second species involved in hybridization with C. calliope was probably C. ob- scura, since it’s the only species of all Calliope genus that has a black breast and black external tail feathers with white bases similar to those of C. pectoralis. Keywords: interspecific hybridization, NUMT, mtDNA, intergenomic recombination, Calliope calliope, C. obscura, C. pectoralis. ГЕНЕТИКА том 55 №1 2019
Вы также можете почитать

















































