Генетика для студентов КРИ - Часть 5. Механизм кроссинговера Анна Михайловна Гусаченко
←
→
Транскрипция содержимого страницы
Если ваш браузер не отображает страницу правильно, пожалуйста, читайте содержимое страницы ниже

Генетика
для студентов КРИ
Электронный лекционный курс – мультимедийная презентация
Анна Михайловна Гусаченко
Часть 5. Механизм кроссинговера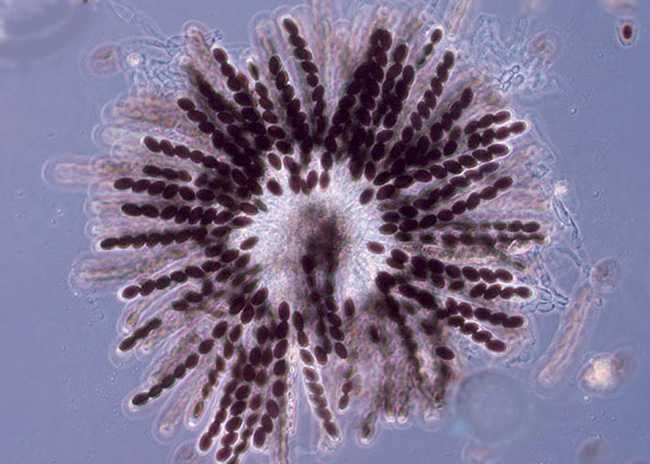
Цитологическое доказательство кроссинговера Эксперимент К. Штерна ♀ ♂ (1930). Хромосомы дрозофилы Некроссоверы кроссоверы маркированы цитологически: одна Х-хромосома несет ♀ ♀ кусочек гетерохроматина от Y, другая Х-хромосома короче из-за транслокации участка на 4 хромосому. Генетические маркеры : cr- глаза цвета гвоздики, cr+ - дикий тип, В – узкие глаза, В+ - нормальные
Цитологическое доказательство кроссинговера Эксперимент Х. Крейтон и Б. МакКлинток на кукурузе. Маркеры цитологические: хромосома имеет блок некроссоверы гетерохроматина и более длинная из-за транслоцированного куска другой хромосомы. Маркеры генетические: кроссоверы с - бесцветный, с+ - окрашенный эндосперм, wx – восковой, wx+ - нормальный эндосперм
Вывод: при кроссинговере хромосомы
обмениваются участками и у животных, и
у растений.
На какой стадии происходит
кроссинговер?
До репликации (2 хроматиды)
или
после репликации
(4 хроматиды)?• Бриджес, Андерсон (1925) показали, что
кроссинговер происходит на стадии 4-х
хроматид у дрозофилы на линии со
сцепленными Х-хромосомами
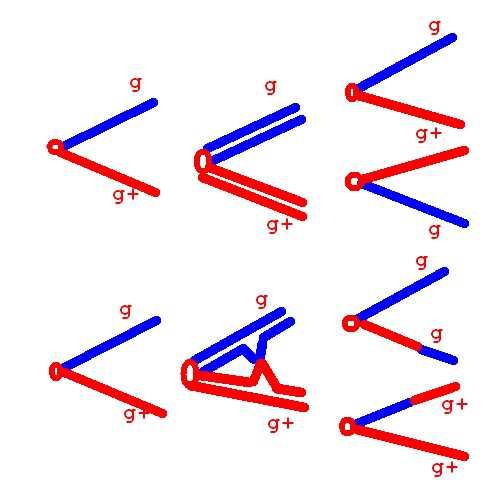
g
g Y x g+
g+ Y
F1 дикий тип дикий тип
g+
Редкие самки- gg -гомозиготыБриджес, Андерсон (1925). Кроссинговер в
сцепленных Х-хромосомах между геном и
центромерой
F1:
Без g/g+
кроссинговера
g/g+
F1:
Кроссинговер
между g и g/g алые
центромерой глаза
g+/g+Обнаружен «митотический»-
соматический кроссинговер
К.Штерн (1936)
Кроссинговер в
соматических
клетках
возникает как
случайное,
мутационное
событие.
Происходит по
Клон singet похожему
механизму, что и
мейотический
на стадии 4-х
хроматид
Клон yellow«Митотический»- соматический
кроссинговер
Два мутантных
клона на
клыле
дрозофилы.
Мутации по
форме
щетинок
Дикий типПлесневый гриб Neurospora crassa
идеален для изучения кроссинговера Цикл развития
Neurospora crassa
Гаплоид, N
Диплоид, 2N, мейоз
Гаплоидные спорыЦикл развития плесени
Neurospora crassa
Аски – «сумки» с гаплоидными спорами.
Каждый аск - продукт одного мейоза –
лежат рядом, удобны для анализаКроссинговер у плесени
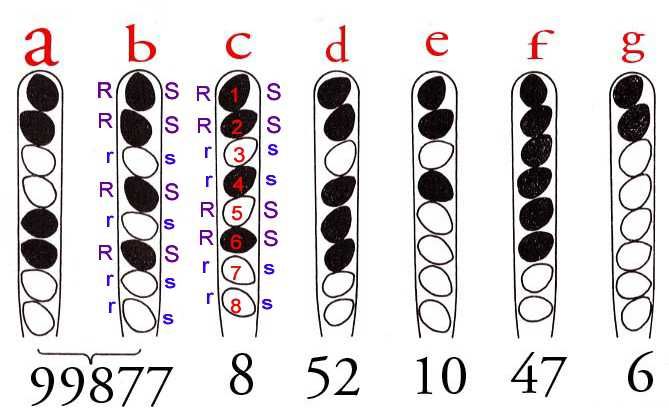
Neurospora crassa
А A
Некроссоверный аск
А
А A
А A
А1 А А2 митоз
A
a a
a
a a
a
a a
Нет кроссинговера между геном и a
центромерой
А A а
Кроссоверные аски
А
A а
А
а а а А
А1 А2 митоз а А
А А а а
а а а
а А А
Кроссинговер между геном и А А
а
центромеройКроссинговер у Neurospora crassa Модель: кроссинговер происходит до репликации на стадии 2-х хроматид Модель: кроссинговер происходит после репликации на стадии 4-х хроматид
Тэйлор (Taylor J.H., 1965) применил новый
метод – радиоавтографию – для
доказательства обмена между хроматидами
• Сперматогенез у кузнечика Romalea microptera.
• Тэйлор вводил кузнечикам тимидин, в котором
вместо Н был его тяжелый изотоп Н3Доказательства обмена между хроматидами
Что такое тритий Н ? 3
• Триитий, сверхтяжёлый водород с молекулярной
массой 3, символы T и 3H — радиоактивный изотоп
водорода. Ядро трития состоит из протона и двух
нейтронов: p+ + 2n0
• В процессе распада тритий превращается в 3He с
испусканием электрона и антинейтрино (бета-
распад), период полураспада — 12,32 года.
Доступная энергия распада очень мала (18,59 кэВ),
средняя энергия электронов 6,5 кэВ.
• Электроны вылетают в разные стороны на
расстояние 1 мкм. Это расстояние удобно для
работы с клетками и хромосомами.Тэйлор (Taylor J.H.) 1965
Доказательства обмена между хроматидами
• Надо, чтобы одна хроматида была меченая, другая
нет. Как это сделать?
• Как регистрировать (увидеть) метку?
Делают препарат хромосом. Используют фотоэмульсию
с AgBr, AgJ. Электроны восстанавливают Ag+ до
металлического серебра Ag0
А1 - М2Тэйлор (Taylor J.H.) 1965
Доказательства обмена между хромосомами
• Надо, чтобы одна хроматида была меченая, другая нет.
• Прошла 1 репликация. Одна хроматида меченая, другая – нет.
• В М2 видны можно увидеть обмены между меченой и
немеченой хроматидой (это половина всех обменов).
• Частота хиазм в самой большой хромосоме 3.67 –
кроссоверных хроматид 3.67/2 = 1.88, мы можем видеть
половину из них: 0.94. Тэйлор увидел 0.89
Подтвердил
гипотезу
«разрыв-
воссоединение»
А1 - М2Генная конверсия у грибов Линдегрен К.К. (1949) открыл необычное расщепление. Нормальное расщепление в тетрадах 4:4 (2:2:2:2). Иногда встречаются 6:2 и даже 3:5. Нарушается закон чистоты гамет. Объяснил с помощью гипотезы Х.Винклера (1930) о генной конверсии
Образование полухиазмы (модель Холлидея),
ее разрезание по конверсионному и
кроссоверному типу (1964 и позже)
конверсия
кроссинговер
Полухиаз
ма
миграция Коррекция
ветвей гетеродуплексаОбразование полухиазмы (модель Холлидея),
ее разрезание по конверсионному и
кроссоверному типу
Полухиазма
в клетках
E. coli
(Potter, конверсия
Dressler, кроссинговер
1976)Образование полухиазмы (модель Холлидея),
ее разрезание по конверсионному и
кроссоверному типу
Полухиазма – хиазма по одной из комплементарных цепей ДНК
Полухиазма может сдвигаться – это миграция ветвей
Полухиазма может изомеризоваться – разворачиваться в крест в
одном и в другом направлении.
Крест полухиазмы может разрезаться двумя способами:
•по той ветви, которая участвовала в полухиазме – это можно
заметить как конверсию,
•по той ветви, которая не участвовала – это будет кроссинговер
В месте кроссинговера или конверсии может возникнуть
гетеродуплекс – участок, где нуклеотиды не комплементарны (это
разные аллели).
Гетеродуплекс корректируется ферментами репарации по любой из
2-х цепей, но уже только по одной в участке кроссинговераСовременная
(самая последняя)
модель
кроссинговера
Двуцепочечный
разрыв:
Режутся оби цепи ДНК
В механизмах
рекомбинации еще
очень много неясного,
это направление
развивается
Кроссинговер, кроссинговера нет,
конверсия (?) конверсияВы также можете почитать

















































