ВЛИЯНИЕ ГОРМОНАЛЬНОГО ФОНА НА ЭКСПРЕССИЮ РЕЦЕПТОРОВ К ПОЛОВЫМ ГОРМОНАМ И МИГРАТОРНОГО ПОТЕНЦИАЛА КЛЕТОК ЭПИТЕЛИЯ И СТРОМЫ ЭНДОМЕТРИЯ МЫШИ IN VITRO
←
→
Транскрипция содержимого страницы
Если ваш браузер не отображает страницу правильно, пожалуйста, читайте содержимое страницы ниже
ОНТОГЕНЕЗ, 2021, том 52, № 6, с. 453–466
БИОЛОГИЯ РАЗВИТИЯ ЖИВОТНЫХ
(БЕСПОЗВОНОЧНЫХ И ПОЗВОНОЧНЫХ)
УДК 57.085.23
ВЛИЯНИЕ ГОРМОНАЛЬНОГО ФОНА НА ЭКСПРЕССИЮ РЕЦЕПТОРОВ
К ПОЛОВЫМ ГОРМОНАМ И МИГРАТОРНОГО ПОТЕНЦИАЛА КЛЕТОК
ЭПИТЕЛИЯ И СТРОМЫ ЭНДОМЕТРИЯ МЫШИ IN VITRO
© 2021 г. Л. Ш. Измайловаa, *, А. А. Гайдамакаa, О. О. Сербинаa, Е. А. Воротелякa
a
Институт биологии развития им. Н.К. Кольцова РАН, ул. Вавилова, 26, Москва, 119334 Россия
*e-mail: luba.ranaway-94@yandex.ru
Поступила в редакцию 25.06.2021 г.
После доработки 22.07.2021 г.
Принята к публикации 27.07.2021 г.
Подготовка к имплантации эмбриона – это комплексный процесс, включающий в себя изменения
как на клеточном, так и на тканевом уровнях в эпителии и строме эндометрия матки. От регуляции
преимплантационных изменений эндометрия и наступления периода рецептивности зависит
успешность прохождения беременности. Моделирование регуляции функционирования эндомет-
рия in vitro позволяет изучить процесс подготовки эндометрия к имплантации эмбриона, что осо-
бенно важно для разработки терапевтических подходов для лечения заболеваний, ассоциирован-
ных с бесплодием. В работе показано влияние стероидных половых гормонов на изменение поляр-
ности эпителия эндометрия in vitro, что также является немаловажным процессом во время
имплантации in vivo. Также описано взаимное влияние эстрогена и прогестерона как основных ре-
гуляторов функционирования эндометрия на экспрессию их рецепторов.
Ключевые слова: эндометрий, эпителий, строма, эпителио-мезенхимный переход, половые стеро-
идные гормоны
DOI: 10.31857/S0475145021060069
ВВЕДЕНИЕ ходимы для исследования раннего постимпланта-
Внутренняя оболочка матки, эндометрий, под- ционного развития. Этот этап мало изучен, так как
держивает имплантацию и дальнейшее развитие прикрепившийся эмбрион развивается внутри
эмбриона. Основные клеточные популяции эндо- матки, что затрудняет наблюдение и манипуляции
метрия представлены клетками люминального и с ним. Для решения этой проблемы использование
железистого эпителия и подлежащей стромы. В хо- мыши в качестве модельного объекта в системах
де приобретения рецептивности эндометрий про- in vitro является перспективным направлением.
ходит серию последовательных изменений. Про- Например, в одной из последних работ было пока-
цесс преобразования эндометрия перед импланта- зано развитие эмбриона мыши in vitro от стадии
цией у человека и мыши управляется двумя гаструляции до позднего органогенеза (Aguilera-
основными гормонами: эстрогеном (E2) и проге- Castrejon et al., 2021). Также для мыши была показа-
стероном (P4), которые активируют соответству- на имплантация эмбриона in vitro с последующим
ющие сигнальные пути (Wang, Dey, 2006). развитием зародышевого цилиндра (Bedzhov et al.,
2014). Работа с мышью помогает избежать этиче-
Нарушение процесса имплантации у человека ских вопросов при исследовании как имплантации
является причиной более чем половины случаев эмбриона и его развития, так и функционирования
потери беременности на ранних сроках (Wilcox эндометрия во время беременности. Модели с ис-
et al., 1988). В связи с этим молекулярная регуля- пользованием клеток эпителия и стромы, выде-
ция имплантации активно изучается. ленных из эндометрия мыши, позволяют рекон-
Изучение имплантации эмбриона человека в струировать имплантацию in vitro.
матке по техническим и этическим причинам за-
труднено, поэтому исследования в этой области Подготовка к имплантации у мыши начинает-
проводятся с использованием модельных живот- ся с секреции преовуляторного Е2 фолликуляр-
ных. Одним из распространенных модельных ными клетками яичников. Функция E2 обеспечи-
объектов является мышь (Mus musculus). Помимо вается путем его взаимодействия с ядерными ре-
решения проблем, возникающих при репродук- цепторами ERα (Estrogen Receptor α) и ERβ
ции, модели имплантации эмбрионов in vitro необ- (Estrogen Receptor β). ERα является доминирую-
453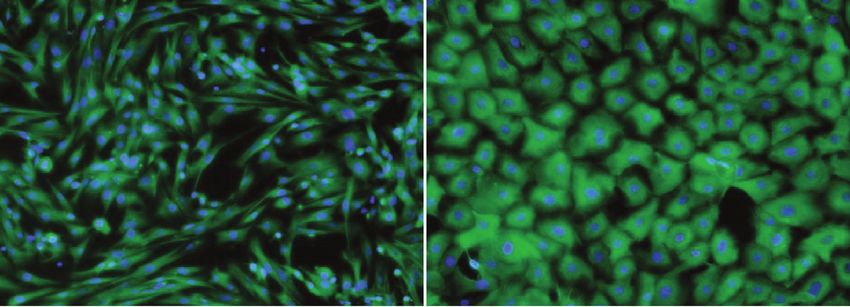
454 ИЗМАЙЛОВА и др.
щей формой рецепторов к E2 в матке мыши, по- ный путь, индуцируемый Р4 через PR необходим
скольку она отвечает за подготовку эндометрия для прохождения имплантации. На этапе, когда
к имплантации (Vasquez and DeMayo, 2013). На полностью сформирована децидуальная зона,
1–2 сутки беременности Е2 запускает пролифера- экспрессия PR в эпителии уже не детектируется. В
цию эпителия, связываясь с ERα стромы (Vasquez то же время у псевдобеременных мышей снижение
and DeMayo, 2013). В строме включается экспрес- уровня экспрессии PR в эпителии происходит
сия инсулин-подобного фактора роста-1 (Insulin- только если искусственно индуцирована дециду-
like growth factor 1; IGF-1) и белков семейства фак- альная реакция. Таким образом, прекращение экс-
торов роста фибробластов (Fibroblast growth fac- прессии PR в эпителии эндометрия регулируется
tors; FGFs), которые паракринно действуют на локальными факторами, продуцируемыми в матке,
эпителиальные клетки и активируют пролифера- и регулируется за счет диалога между стромальным
цию (Hantak et al., 2014). Кроме того, фоллику- и эпителиальным компартментами (Honglu et al.,
лярный Е2 обеспечивает подготовку клеток стро- 2012).
мы к децидуализации и активирует экспрессию
Для создания адекватной модели эндометрия
рецепторов к P4 (Progesterone receptors; PRs) в эн-
необходимо понимание изменений, происходя-
дометрии. Также были получены данные, не со-
щих в ответ на гормональную стимуляцию в на-
гласующиеся с приведенными выше результата-
тивных клетках эпителия и стромы. Регуляция
ми. Тан с соавторами показали, что в первый день
экспрессии рецепторов к половым гормонам в те-
беременности ER экспрессируется только в эпи-
чение эстрального цикла и беременности, а также
телии, но не в строме (Tan et al., 1999). Однако,
роли взаимодействия эпителия и стромы требует
эти данные были получены методом FISH с де-
отдельного изучения. Также исследования в этой
текцией при помощи метода авторадиографии,
области помогут расширить знания о регуляции
что затрудняет однозначную трактовку получен-
эндометрия in vivo.
ных результатов. К 4 суткам, когда желтое тело
уже сформировано, наблюдается небольшой пик Люминальный эпителий (ЛЭ) эндометрия яв-
E2, который участвует в активации эмбриона и ляется первым барьером на пути инвазии эмбри-
подготовке стромы к децидуальной реакции она. Эпителий в ходе имплантации претерпевает
(Zhang et al., 2013). Растущий на 3 сутки беремен- трансформацию цитоплазматической мембраны
ности уровень P4 вызывает в эндометрии измене- (ТЦМ), которая характеризуется изменением по-
ния, необходимые для поддержания развития ляризации клеток. Этот процесс начинается перед
прикрепившегося эмбриона. P4 действует через инвазией бластоцисты и, вероятно, необходим для
ядерные рецепторы – PRs. Они также имеют 2 облегчения прохождения эмбриона сквозь эпите-
изоформы: PRA и PRB, которые транскрибиру- лиальный барьер. Во многом ТЦМ сходна с эпите-
ются под разными промоторами с одного гена лио-мезенхимным переходом (ЭМП), свойствен-
(Vasquez and DeMayo, 2013). P4 подавляет экс- ным опухолевым клеткам.
прессию ERα в эндометрии, пролиферацию эпи-
Ряд авторов высказали предположение, что в
телиальных клеток и, наоборот, активирует про-
околоимплантационный период ЛЭ проходит
лиферацию клеток стромы, которая связана с де-
именно ЭМП и приобретает способность к ми-
цидуальной реакцией (Zhang et al., 2013). В
грации. Эта гипотеза была проверена на линии
дополнение к этому, Р4 вызывает ремоделирова-
клеток аденокарциномы эндометрия (Ishikawa –
ние эпителиального барьера, которое необходимо
Ишикава). Ишикава – это клеточная линия, ис-
для облегчения прикрепления и инвазии эмбриона
пользуемая в качестве модели рецептивного эпи-
(Ye, 2020). Исходя и этих данных, можно заклю-
телия эндометрия. В работе Учиды с соавторами
чить, что баланс эстроген-зависимых и прогесте-
(Uchida et al., 2012) показали, что культивирование
рон-зависимых эффектов во многом обуславлива-
клеток эпителия эндометрия в среде с E2 и P4 вы-
ет успешное прохождение имплантации.
зывает повышение миграторного потенциала кле-
Несмотря на то, что существует немало работ в ток, перестройку актинового цитоскелета с образо-
этой области, до сих пор не выяснено, каким об- ванием стресс-фибрилл актина и повышение уров-
разом половые стероидные гормоны влияют на ня экспрессии виментина. Также в эпителиальных
экспрессию их рецепторов в эпителиальном и клетках как при обработке гормонами, так и при
стромальном компартментах эндометрия во вре- добавлении сфероидов из клеток хориокарцино-
мя беременности. Также не ясно, вносит ли взаи- мы человека (JAR-cells) происходило характерное
модействие клеточных популяций эпителия и для ЭМП “кадгериновое переключение”: сниже-
стромы какой-либо вклад в изменение экспрес- ние уровня экспрессии Е-кадгерина и повыше-
сии рецепторов к половым гормонам. Например, ние уровня экспрессии N-кадгерина. Монослой-
для мыши было показано, что PR экспрессирует- ная культура клеток Ишикава поддерживала при-
ся в ЛЭ в околоимплантационные сроки, уровень крепление сфероидов in vitro, при этом площадь
его экспрессии снижается с началом децидуали- разрастания сфероидов была больше, если клетки
зации стромы. При этом, показано, что сигналь- предварительно культивировали в среде с E2 и P4.
ОНТОГЕНЕЗ том 52 №6 2021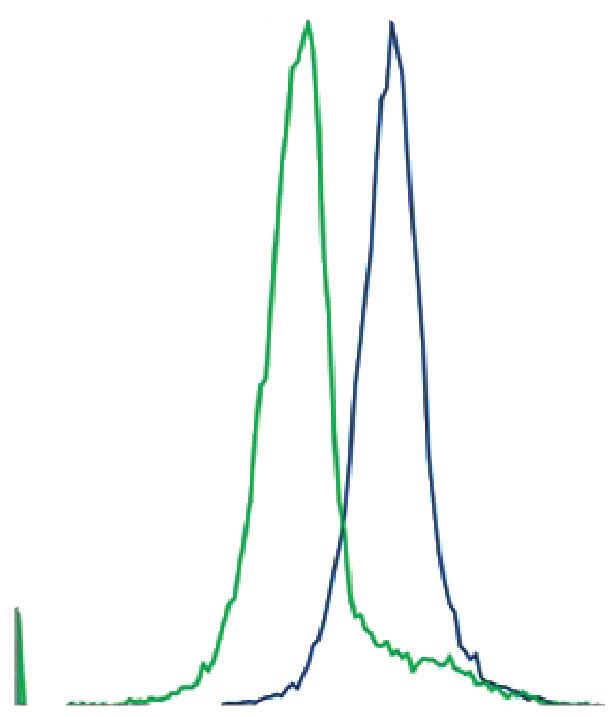
ВЛИЯНИЕ ГОРМОНАЛЬНОГО ФОНА НА ЭКСПРЕССИЮ РЕЦЕПТОРОВ 455
Авторы предложили модель, согласно которой шивании. Далее отбирали и центрифугировали
клетки ЛЭ приобретают способность к миграции раствор с клетками стромы эндометрия. Строму
и перемещаются по направлению от зоны им- промывали полной средой ДМЕМ и осаждали
плантации, тем самым облегчая прохождение эм- центрифугированием. Осадок с клетками эпите-
бриона сквозь эпителиальный барьер (Uchida лия или стромы переносили в лунки 12–96 луноч-
et al., 2012). Однако этот результат получен с ис- ного планшета или чашки Петри.
пользованием опухолевых клеток.
Целью данной работы было проверить гипоте-
зу о прохождении клетками эпителия ЭМП in vitro и Культивирование клеток эпителия
внести ясность в вопрос о влиянии половых сте- и стромы эндометрия
роидов на уровень экспрессии их рецепторов в Клетки эпителия и стромы культивировали в
клетках эпителия и стромы матки мыши in vitro. полной минимальной среде Игла ДМЕМ в инку-
баторе в стандартных условиях – 37°С, 5% СО2.
МАТЕРИАЛЫ И МЕТОДЫ Полная среда ДМЕМ состояла из среды ДМЕМ
(Gibco, Ирландия), 10% эмбриональной телячьей
Животные сыворотки (ЭТС), 1% Glutamax (Gibco, Ирландия),
В работе были использованы самки мыши ли- 1% PenStrep (Gibco, Ирландия). Для моделирова-
нии Balb/C в возрасте 8–12 недель. Животные со- ния гормонального воздействия на эпителий и
держались в стандартных условиях вивария ИБР строму эндометрия, соответствующего имплан-
РАН по 4–6 особей в клетке, со свободным досту- тации, в среду добавляли гормоны, разведенные в
пом к корму и воде. В каждом эксперименте было диметилсульфоксиде (DMSO; SigmaAldrich, США)
задействовано по 6 особей, которые до начала с конечной концентрацией Е2 – 0.9 нМ и Р4 –
эксперимента содержались в одной клетке. Все 63.5 нМ (SigmaAldrich, США). Конечная концен-
манипуляции с животными были одобрены эти- трация ДМСО (SigmaAldrich, США) в среде после
ческой комиссией ИБР РАН (Протокол № 40 от добавления в нее гормонов не превышала 0.02%.
17.09.2021). Культивирование клеток проводили в 4-х гормо-
нальных режимах – контроль без добавления гор-
Для получения культур клеток эпителия и монов, с добавлением Е2, с добавлением Р4 и с обо-
стромы матки самок мыши стимулировали E2 для ими гормонами. Концентрации гормонов, соот-
синхронизации в фазе эструса по стандартному ветствующие гормональному фону мыши в период
протоколу (De Clercq et al., 2017). имплантации, были подобраны по результатам
анализа литературы (Ye et al., 2012). Смена сред
Получение культур клеток эпителия проводилась каждые 1–2 дня.
и стромы матки
Для получения культур клеток животных вы- Иммуноцитохимия
водили из эксперимента при помощи цервикаль-
ной дислокации и изолировали матки. Рога маток Для иммуноцитохимического окрашивания
промывали раствором Хенкса (ПанЭко, Россия) с клетки предварительно были посажены в лунки
добавлением 1% раствора гентамицина и разреза- 48 луночного планшета. Фиксацию, отмывки и
ли продольно. Из ткани маток всех животных, окраску первичными и вторичными антителами
участвовавших в эксперименте, выделяли тоталь- (табл. 1) проводили по стандартному протоколу
ные фракции клеток эпителия и стромы, которые (De Clercq et al., 2017). Клетки анализировали при
затем распределялись по группам в зависимости помощи флуоресцентного микроскопа Olympus
от времени культивирования и типа гормональ- IX51 (Olympus, Япония). В качестве контроля ис-
ного воздействия. Для выделения эпителия ткань пользовали образцы клеток, окрашенных без до-
рогов матки помещали в 0.1% раствор диспазы бавления первых антител.
при 4°С на 12 ч. На следующий день ткань в рас-
творе диспазы (Gibco, Ирландия) инкубировали Проточная цитометрия
при 37°С в течение 15 мин с последующим интен-
сивным встряхиванием на вортексе в течение Клетки снимали с пластика стандартным спо-
2 мин. Среду с отделившимися эпителиальными собом (Kubaczka et al., 2014). Пробоподготовку
пластами отбирали и центрифугировали. Эпите- для анализа клеток при помощи метода проточ-
лий промывали полной средой ДМЕМ, осажда- ной цитофлуориметрии проводили стандартным
ли центрифугированием. Для выделения стромы способом (Shakya et al., 2020). Образцы анализи-
оставшуюся после выделения эпителия ткань ровали на проточном цитофлуориметре Attune
матки гомогенизировали и помещали в 0.2% NxT (ThermoFisher, США). В качестве контроля
раствор коллагеназы первого типа (Gibco, Ир- использовали образцы клеток, окрашенных без
ландия) на 5 ч при 37°С при постоянном переме- добавления первичных антител.
ОНТОГЕНЕЗ том 52 №6 2021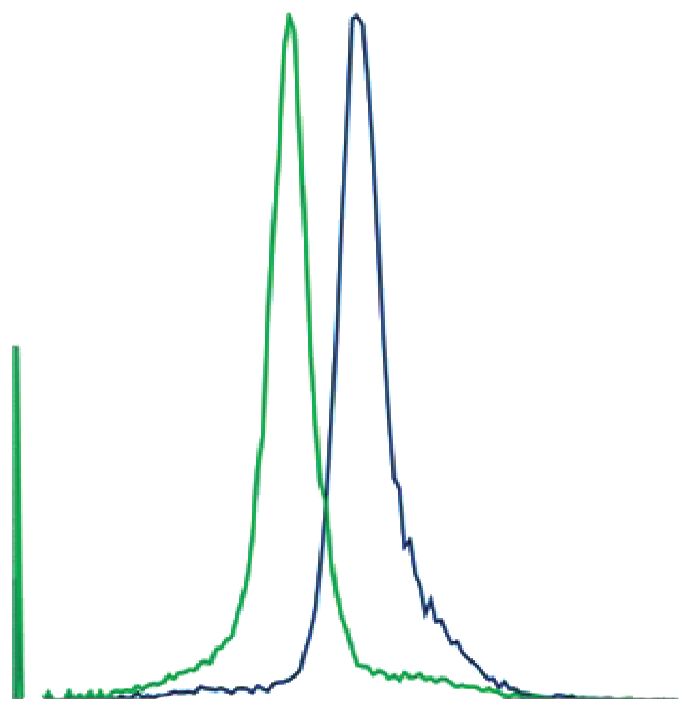
456 ИЗМАЙЛОВА и др.
Таблица 1. Список антител, использованных в работе
Название В каком животном изготовлено Фирма, каталожный номер
I Polyclonal Anti-Vimentin antibody Chicken Abcam ab24525
I Monoclonal Anti-PanCytokeratin antibody Mouse Abcam ab7753
II Anti-Сhicken Alexa-488 conjugated antibody Goat Thermo Fisher A11039
II Anti-Mouse Alexa-488 conjugated antibody Goat Thermo Fisher A32723
Определение времени удвоения культуры реальном времени с применением смеси для ПЦР
qPCRmix-HS SYBR (Евроген, Россия) по ин-
Время удвоения культуры определяли на при- струкциям производителя на приборе LightCycler 96
боре xCELLigence (RTCA, CША) стандартным (Roche, Швейцария). В качестве контроля в каж-
способом (Hamidi et al., 2017). дом повторе эксперимента для каждого праймера
(табл. 2) были проанализированы пробы RT–, то
ПЦР-РВ (полимеразная цепная реакция есть пробы, в которые на этапе обратной тран-
с детекцией в реальном времени) скрипции не была добавлена обратная тран-
скриптаза. Данные пробы не содержали к-ДНК и
Из клеток эпителия, стромы эндометрия мы- их анализ методом ПЦР-РВ был показателем ге-
ши или гомогената ткани матки мыши выделяли номной ДНК в пробе. Полученные результаты
тотальную РНК с помощью Quick-RNA Miniprep были нормированы на экспрессию референсного
Kit (Zymo Research, США) по инструкциям про- гена ActinB, выбранного на основании обзора ли-
изводителя. Затем синтезировали кДНК с помо- тературы (Lin et al., 2013).
щью набора для обратной транскрипции MMLV
RT kit (Евроген, Россия) со случайными прайме-
рами по инструкциям производителя. После это- Модель раны и оценка уровня миграции клеток
го проводили количественную ПЦР с детекцией в эпителия эндометрия
Клетки эпителия эндометрия культивировали
при каждом гормональном режиме 2 сут до дости-
Таблица 2. Праймеры, использованные при проведе-
нии ПЦР-РВ. Все праймеры были произведены фир-
жения 90% конфлюэнтности. Затем микропипет-
мой Евроген, Россия кой с пластиковым носиком удаляли клетки по-
лосой в монослое, анализировали и фотографи-
Название Последовательность ровали область раны. Через 24 ч после нанесения
moVim-F GCCAGGCCAAGCAGGAGTCAAAC раны ее повторно фотографировали (рис. 1). Ана-
лиз зарастания раны производился описанным
moVim-R ATCCTGCAGGCGGCCAATAGTGT ранее методом (Uchida et al., 2012).
moCdh1-F GCTGGACCGAGAGAGTTACC
moCdh1-R CCGGGCATTGACCTCATTCT
Статистические методы
moKrt8-F AACAAGTTCGCCTCCTTCATT
moKrt8-R CATGTTGTCCATGTTGCTCCTC Данные анализировали с помощью Microsoft
Excel и программы GraphPad Prism 5.0 (GraphPad
moPgr-F CAATGGAAGGGCAGCATAACT Software, v.5, США). Для статистического анализа
moPgr-R ACACCGTCAAGGGTTCTCATA данных и построения графиков использовали
moEsr1-F AGGCTTTGGGGACTTGAATCT программу GraphPad® Prism (GraphPad Software,
moEsr1-R CATGCGGAACCGACTTGAC v.5, США). Если распределение сравниваемых
значений параметра в нескольких независимых
moActb-F ACCCGCCACCAGTTCG выборках отличалось от нормального, то для ста-
moActb-R AGCATCGTCGCCCGC тистической обработки результатов использова-
moTbp-F ACCGTGAATCTTGGCTGTAAA ли непараметрический аналог ANOVA – ранго-
moTbp-R GTCTGGATTGTTCTTCACTCTTG вый дисперсионный анализ Краскела–Уоллиса.
Для распределений, не отличающихся от нор-
moCdh2-F GCCCGGTTTCACTTGAGAG мального, использовали дисперсионный анализ
moCdh2-R GTACCGCAGCATTCCATTCA ANOVA. Проверку гипотезы о нормальности рас-
moSnai2-F GCCTCCAAGAAGCCCAACTA пределения проводили с помощью критерия Кол-
moSnai2-R GCCGACGATGTCCATACAGT могорова–Смирнова. Статистически значимыми
приняты данные со значением P < 0.05. Для по-
moTwist2-F GGACCAAGGCTCTCAGAACAA парного сравнения двух нормально распределен-
moTwist2-R TCCAGGCTTCCTCGAAACAG ных зависимых выборок использовался парный
ОНТОГЕНЕЗ том 52 №6 2021ВЛИЯНИЕ ГОРМОНАЛЬНОГО ФОНА НА ЭКСПРЕССИЮ РЕЦЕПТОРОВ 457
(а) (б)
Рис. 1. Оценка клеточной подвижности. (а) – область раны сразу после ее нанесения; (б) – область раны через 24 часа
после нанесения.
t-критерий Стьюдента. Для выборок, имеющих Эти эксперименты подтвердили эпителиальную
не нормальное распределение, использовали не- природу культуры эпителия эндометрия и мезен-
параметрический аналог – критерий Уилкоксона. химальную природу культуры клеток стромы эндо-
Для сравнения двух несвязанных выборок приме- метрия мыши, что демонстрирует эффективность
нялся t-критерий Стьюдента для независимых протокола их выделения и чистоту полученных
выборок и его непараметрический аналог – кри- культур. Для дальнейшей характеризации выде-
терий Манна–Уитни. ленных культур клеток скорость их пролиферации
была оценена при помощи прибора xCELLegence
(RTCA). Среднее время удвоения популяции кле-
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ ток эпителия составляло 100 ч и было достоверно
Характеристика полученных культур выше, чем среднее время удвоения клеток культу-
ры стромы эндометрия мыши (69 ч).
Культуры эпителия и стромы выделяли из эн-
дометрия матки мыши по разработанному нами
протоколу: подобрали время и условия инкуби- Уровень миграции клеток эпителия эндометрия
рования с растворами ферментов (см. Материалы при разных гормональных
и методы), благодаря которым в рамках одного режимах культивирования
эксперимента возможно выделить из матки мы- Для оценки влияния половых стероидных гор-
ши и эпителий, и клетки стромы. Это позволяет монов на способность к миграции эпителия эн-
использовать культуры клеток эпителия и стромы дометрия была исследована подвижность клеток
от одной группы животных, что повышает досто- на модели раны. Через 24 ч после нанесения цара-
верность экспериментов. пины были обнаружены статистически значимые
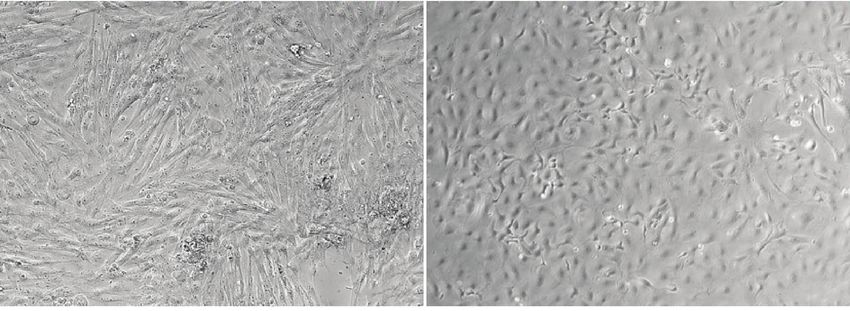
Стромальные и эпителиальные клетки в культу- различия в изменении площади раны практиче-
ре имели фибробластоподобную и эпителиальную ски между всеми группами, кроме образцов, об-
морфологию, соответственно (рис. 2в, 2г). Клеточ- работанных обоими гормонами (E2P4) и контро-
ные культуры стромы и эпителия были окрашены лем (без обработки гормонами; Б/Г) (рис. 4). При
на мезенхимальные (виментин) (рис. 2а, 2д) и эпи- этом степень закрытия раны была наибольшей в
телиальные (PanCK) (рис. 2б, 2е) маркеры и про- группе клеток, обработанных Р4, что говорит о
анализированы при помощи методов иммуноци- высокой клеточной подвижности. Самая низкая
тохимии и проточной цитофлуориметрии. подвижность клеток была обнаружена в культуре,
Была проведена оценка экспрессии эпители- которая находилась в среде с Е2. Уровень мигра-
альных и мезенхимальных маркеров в клетках ции в группе клеток, обработанных обоими гор-
эпителия и стромы при помощи метода ПЦР-РВ монами, имел промежуточные значения.
(рис. 3). В этом эксперименте в качестве эпители- В ходе анализа уровня миграции опухолевых
альных маркеров использовали Е-кадгерин и спе- клеток в модели раны было обнаружено досто-
цифический маркер эпителия эндометрия кера- верное повышение подвижности клеток при
тин 8. Для оценки уровня экспрессии стромаль- культивировании в среде с обоими гормонами
ных маркеров использовали виментин. (Uchida et al., 2012). Мы также наблюдали увели-
ОНТОГЕНЕЗ том 52 №6 2021458 ИЗМАЙЛОВА и др.
(а) (б)
100 мкм 100 мкм
(в) (г)
100 мкм 100 мкм
(д) (е)
100 100
75 75
Percent of Max
Percent of Max
50 50
25 25
0 0
100 101 102 103 104 105 106 100 101 102 103 104 105 106
Рис. 2. Культуры клеток, выделенные из эндометрия мыши. Иммуноцитохимическая окраска клеток, (а) строма,
окраска антителами к виментину и DAPI, (б) эпителий, окраска антителами к PanCK и DAPI. Общий вид культуры,
фазовый контраст, (в) строма, (г) эпителий. Проточная цитометрия, (д) строма, окраска антителами к виментину,
(е) эпителий, окраска антителами к PanCK. На графиках представлено сравнение интенсивности флуоресценции
окрашенных образцов (синий) и контроля (зеленый).
чение уровня миграции клеток в условиях куль- наименьшая – в среде с E2. Исходя из получен-
тивирования с Е2Р4, однако только относительно ных данных, мы пришли к выводу, что Р4 активи-
обработанных Е2 клеток, но не контрольной рует способность к миграции эпителия, а Е2 –
группы. При этом наибольшая подвижность кле- подавляет ее. Известно, что Е2 оказывает эффект,
ток обнаружена в культуре, обработанной P4, а поддерживающий полярность эпителия, и вос-
ОНТОГЕНЕЗ том 52 №6 2021ВЛИЯНИЕ ГОРМОНАЛЬНОГО ФОНА НА ЭКСПРЕССИЮ РЕЦЕПТОРОВ 459
1.5 Относительный уровень миграции клеток
эпителия в зависимости от действия
гормонов
1 500 000
Изменение площади раны
1.0
(Delta), мкм
1 000 000
0.5
500 000
0
im
r t8
ad
m
r t8
d
ca
Vi
-c
V
аK
K
Е-
аЕ 0
ма
й
ий
ли
ом
ий
о
ом
ел
те
тр
тр
Г
E2
P4
P4
ел
ит
тр
Б/
и
С
С
ит
E2
Эп
Эп
С
Эп
Тип гормонального воздействия
Рис. 3. Анализ экспрессии генов эпителиальных и ме- Рис. 4. График зависимости уровня миграции клеток
зенхимальных маркеров в культурах клеток эпителия эпителия эндометрия от типа гормонального воздей-
и стромы, выделенных из эндометрия матки. Уровень ствия. Статистически значимые различия обнаруже-
экспрессии нормализован на экспрессию рефе- ны между всеми группами, кроме Е2Р4 и Б/Г (без об-
ренсного гена ActinB. работки гормонами). P-значение ≤0.006.
станавливает микроворсинки на апикальной по- На первом этапе была исследована экспрес-
верхности клеток (Quinn et al., 2019; Whitby et al., сия генов Cdh1 и Cdh2, кодирующих Е-кадгерин
2020). Следовательно, подавление миграции кле- и N-кадгерин, соответственно (рис. 5). Экспрес-
ток эпителия эндометрия может быть связано с сия эпителиального маркера, Cdh1, достоверно
ингибирующим действием Е2. Необходимо также уменьшалась в эпителии к 5 сутками культиви-
подчеркнуть, что именно Р4 является основным рования (p < 0.01). В то же время экспрессия мезен-
регулятором процесса имплантации. Кроме того, химного маркера, Cdh2, увеличивалась (p < 0.01).
показана роль контролируемых Р4 факторов в Однако уровень экспрессии этих генов не зависел
снижении апико-базальной полярности эпите- от типа гормонального воздействия.
лия эндометрия (Zhang et al., 2013; Kallen et al., На втором этапе была изучена экспрессия сле-
2014). Этот результат согласуется с данными, по- дующей пары генов: Krt8 и Vim (рис. 6). Эти гены
лученными в более ранних исследованиях, и может также кодируют маркеры эпителия (кератин-8) и
частично подтверждать предположения о приобре- мезенхимы (виментин). В отличие от Cdh1 экс-
тении подвижности клетками люминального эпи- прессия Krt8 в целом не зависела от времени куль-
телия во время имплантации in vivo и ключевом тивирования. При этом экспрессия Vim анало-
влиянии Р4 на этот процесс. Поэтому наша модель гично Cdh2 достоверно увеличивалась на 5 сут
в контексте оценки подвижности клеток является культивирования (p < 0.01). Достоверных разли-
релевантной и отражает некоторые гормон-зависи- чий в экспрессии этих генов между типами гор-
мые эффекты, действующие в ходе имплантации. мональных режимов культивирования не наблю-
далось.
Уровень экспрессии мезенхимальных, Далее была проведена оценка уровней экс-
эпителиальных маркеров и генов-маркеров ЭМП прессии генов Twist2 и Snai2, кодирующих факто-
в культуре эпителиальных клеток эндометрия ры транскрипции ЭМП (рис. 7). Их экспрессия
при различных гормональных режимах также не зависела от типа гормонального воздей-
культивирования ствия, при этом уровень Snai2 достоверно увели-
Для оценки влияния гормонов на изменение чивался к 5 дню культивирования (p < 0.01), а
эпителиального фенотипа клеток эпителия эндо- Twist2 – практически не менялся со временем.
метрия со временем культивирования был прове- Процесс ТЦМ сходен с эпителио-мезенхим-
ден ПЦР-РВ генов, кодирующих маркеры эпите- ным переходом (ЭМП), свойственным опухоле-
лия, мезенхимы и факторы транскрипции ЭМП. В вым клеткам. Например, в ходе обоих процессов
качестве образцов для анализа экспрессии вы- происходит разборка фокальных контактов, что
бранных генов были использованы культуры эпи- способствует индивидуализации клеток (Lam-
телия эндометрия, которые находились в средах с ouille et al., 2014; Kaneko et al., 2011, 2011). Также
гормонами в течение 2 и 5 сут. апикальная поверхность эпителиальных клеток
ОНТОГЕНЕЗ том 52 №6 2021460 ИЗМАЙЛОВА и др.
Cdh1 Cdh2
0.15 0.015
0.10 0.010
rQ
0.05 0.005
0 0
5 я
E2 ей
5 я
P4 ей
5 я
P ей
5 я
ей
5 я
E2 ей
5 я
P4 ей
5 я
P ей
5 я
ей
Б / 2 дн
E2 дн
P4 дн
P4 дн
Б / 2 дн
E2 дн
P4 дн
P4 дн
дн
дн
E2 дн
дн
дн
дн
E2 дн
дн
2
2
T2 4 2
2
2
T2 4 2
Г
Г
Г
Г
Б/
Б/
Тип гормонального воздействия Тип гормонального воздействия
Рис. 5. Анализ экспрессии генов эпителиального (Cdh1) и мезенхимального (Cdh2) типов кадгеринов в культурах кле-
ток эпителия, выделенных из эндометрия матки и культивируемых при разных гормональных режимах в течение 2 и
5 сут. Уровень экспрессии нормализован на экспрессию референсного гена ActinB.
Krt8 Vim
2.5 1.0
2.0 0.8
1.5 0.6
rQ
1.0 0.4
0.5 0.2
0 0
я
ей
я
ей
я
P ей
5 я
ей
я
ей
я
ей
я
P ей
5 я
ей
дн
дн
дн
P4 дн
дн
дн
дн
P4 дн
дн
дн
дн
дн
дн
дн
дн
дн
2
2
2
T2 4 2
2
2
2
T2 4 2
5
5
5
5
5
5
Г
E2
P4
Г
E2
P4
Г
E2
P4
Г
E2
P4
Б/
Б/
Б/
Б/
E2
E2
Тип гормонального воздействия Тип гормонального воздействия
Рис. 6. Анализ экспрессии генов эпителиальных (Krt8) и мезенхимальных (Vim) маркеров в культурах клеток эпителия,
выделенных из эндометрия матки и культивируемых при разных гормональных режимах в течение 2 и 5 сут. Уровень
экспрессии нормализован на экспрессию референсного гена ActinB.
теряет микроворсинки (Chávez et al., 1985). В ре- окрашивании были преимущественно локализова-
моделировании актинового цитоскелета как при ны в эпителии эндометрия (Du et al., 2009; Gou et
ЭМП, так и при ТЦМ участвуют малые ГТФазы al., 2019).
семейства RhoA, в дополнение к этому происхо- Однако существует ряд отличий между ЭМП и
дит диссоциация актина от белкового комплекса ТЦМ. При ЭМП происходит разборка плотных
ERM (Ezrin, Radixin, Moesin), связывающего ак- контактов между клетками эпителия, что облег-
тин с белками цитоплазматической мембраны. чает их миграцию. При подготовке к окну им-
Весомым аргументом в пользу гипотезы о ЭМП плантации плотные соединения, наоборот, выяв-
ЛЭ является результат, полученный в работах по ляются на латеральных мембранах клеток ЛЭ эн-
анализу экспрессии генов, которые кодируют фак- дометрия, также присутствуют десмосомы, хотя и
торы транскрипции ЭМП. Экспрессия генов Snai2 в меньшем количестве, чем до наступления ре-
и Twist2 увеличивается в эндометрии беременных цептивности эпителия (Illingworth et al., 2000). В
мышей к моменту имплантации, при этом их бел- то же время при ЭМП белковые комплексы де-
ковые продукты при иммуногистохимическом смосом полностью разбираются для обеспечения
ОНТОГЕНЕЗ том 52 №6 2021ВЛИЯНИЕ ГОРМОНАЛЬНОГО ФОНА НА ЭКСПРЕССИЮ РЕЦЕПТОРОВ 461
Twist2 Snai2
0.0002 0.005
0.0001 0.004
0.003
rQ
0.0001
0.002
0
0.001
0 0
5 я
E2 ей
5 я
P4 ей
5 я
P ей
5 я
ей
5 я
E2 ей
5 я
P4 ей
5 я
P ей
5 я
ей
Б / 2 дн
E2 дн
P4 дн
P4 дн
Б / 2 дн
E2 дн
P4 дн
P4 дн
дн
дн
E2 дн
дн
дн
дн
E2 дн
дн
2
2
T2 4 2
2
2
T2 4 2
Г
Г
Г
Г
Б/
Б/
Тип гормонального воздействия Тип гормонального воздействия
Рис. 7. Анализ экспрессии генов маркеров ЭМП (Twist2, Snai2) в культурах клеток эпителия, выделенных из эндомет-
рия матки и культивируемых при разных гормональных режимах в течение 2 и 5 сут. Уровень экспрессии нормализо-
ван на экспрессию референсного гена ActinB.
миграции клеток (Whitby et al., 2020). На поверх- нов ЭМП не показано в нашей работе. Однако в
ности опухолевых клеток в процессе ЭМП обра- исследовании на клетках аденокарциномы эндо-
зуются небольшие выпячивания цитоплазмати- метрия человека были выявлены гормон-зависи-
ческой мембраны – блебы, а клетки эндометрия мые эффекты на культуру. Так, было выявлено,
во время имплантации формируют пиноподии. что клетки Ишикава в среде с обоими гормонами
Несмотря на то, что обе эти структуры – это бога-
тые актином выступы на поверхности клеток, они проходят “кадгериновое переключение”, форми-
имеют некоторые различия. Блебы участвуют в руют стресс-фибриллы актина, приобретают спо-
экзоцитозе везикул, несущих металлопротеина- собность к миграции и начинают экспрессиро-
зы, тогда как на поверхности пиноподий проис- вать виментин (Uchida et al., 2012). Несмотря на
ходит экзоцитоз LIF (Leukemia Inhibitory Factor), то, что авторы работы приводят аналогии с про-
что способствует имплантации. Таким образом, цессами, происходящими перед инвазией, мы
природа процесса ТЦМ и его роль в имплантации считаем, что поддержание клеток Ишикава в сре-
не до конца выяснены. де с эстрадиолом активирует ER-зависимый путь
Исходя из полученных нами результатов, мож- ЭМП, не связанный с имплантацией (Liu et al.,
но заключить, что клетки эпителия в ходе культи- 2016). Следовательно, клетки Ишикава могут реа-
вирования могут претерпевать “кадгериновое пе- гировать на действие гормонов не как нативные
реключение”, что свойственно для проходящих клетки люминального эпителия эндометрия во
ЭМП клеток. При этом клетки не теряют эпители- время имплантации, а как опухолевые клетки эн-
альные свойства, что отражается в поддержании дометрия на стимулирующие ЭМП сигналы. Из
экспрессии Krt8 на высоком уровне. Мы предпола-
этого можно заключить, что использование опу-
гаем, что эпителий в условиях 2D культуры может
отчасти терять эпителиальный фенотип за счет из- холевых клеток для исследования влияния гормо-
мененной морфологии и поляризации клеток, нов на полярность эпителия эндометрия может
перераспределения межклеточных соединений. привести к получению ложноположительного ре-
Похожий результат был получен в 2D культуре зультата. Наша модель также имеет ограничения,
клеток ретинального эпителия, который после так как условия культивирования могут нести
первого пассажа приобретает черты мезенхимных преобладающий эффект на изменение поляриза-
клеток (Зурина, 2017; Tian et al., 2018). Вероятно, в ции клеток эпителия, чем гормональные воздей-
условиях 2D культивирования эпителий эндо- ствия. Для более точного изучения влияния поло-
метрия может проходить частичный ЭМП, при
котором клетки могут приобретать мезенхимные вых гормонов на клетки эпителия эндометрия не-
маркеры без потери эпителиальных свойств (Nie- обходимо создание 3D систем культивирования,
to et al., 2016). Важно отметить, что влияние поло- например, органоидов, где эпителиальные клетки
вых стероидных гормонов на изменение экспрес- сохраняют свою нативную морфологию в услови-
сии генов-маркеров эпителия, мезенхимы и ге- ях, приближенных к in vivo (Luddi et al., 2020).
ОНТОГЕНЕЗ том 52 №6 2021462 ИЗМАЙЛОВА и др.
Эпителий Esr1 Строма Esr1
0.0004
0.0002
0.0003 0.0002
0.0001
rQ
0.0002
0.0001
0.0001
0
0 0
5 я
E2 ей
5 я
P4 ей
5 я
P ей
5 я
ей
5 я
E2 ей
5 я
P4 ей
5 я
P ей
5 я
ей
Б / 2 дн
E2 дн
P4 дн
P4 дн
Б / 2 дн
E2 дн
P4 дн
P4 дн
дн
дн
E2 дн
дн
дн
дн
E2 дн
дн
2
2
T2 4 2
2
2
T2 4 2
Г
Г
Г
Г
Б/
Б/
Тип гормонального воздействия Тип гормонального воздействия
Рис. 8. Анализ экспрессии гена рецептора к эстрогену в культурах клеток эпителия и стромы, выделенных из эндомет-
рия матки и культивируемых при разных гормональных режимах в течение 2 и 5 сут. Уровень экспрессии нормализо-
ван на экспрессию референсного гена ActinB.
Уровень экспрессии рецепторов к эстрогену увеличением секреции P4 перед имплантацией,
и прогестерону в культурах клеток эпителия что необходимо для снижения E2-зависимой
и стромы эндометрия при различных пролиферации и снижения полярности клеток
гормональных режимах люминального эпителия (Zhang et al., 2013). Так-
Гормональная стимуляция мышей вызывала же мы показали, что процесс Р4-зависимого ин-
наступление фазы эструса, во время которой про- гибирования экспрессии ER не зависел от стро-
исходит оплодотворение и наступает беремен- мы и происходил в эпителии, культивируемом от-
ность. Далее выделенные клетки содержались при дельно от стромальных клеток.
гормональных режимах, в которых, помимо кон- Экспрессия Esr1 в строме не изменялась со
трольной группы, клетки подвергались воздей- временем культивирования, также не было выяв-
ствию гормонов в концентрациях, соответствую- лено гормон-зависимых эффектов (рис. 8). По
щих “окну имплантации” (Ye et al., 2012). имеющимся данным, в начале беременности уро-
Уровень экспрессии Esr1 в клетках эпителия вень экспрессии ER возрастает на 3–4 день в под-
эндометрия снижался во всех эксперименталь- эпителиальном регионе стромы, что предполага-
ных группах со временем культивирования. При ет влияние эпителия на этот процесс. Дальней-
этом, гормон-зависимые отличия между экспе- шие изменения уровня экспрессии ER в строме
риментальными группами клеток эпителия были происходят на 5–7 сут и связаны с образованием
заметны только на второй день культивирования первичной и вторичной децидуальных зон (Tan
и практически полностью нивелировались к 5 дню. et al., 1999). С этим согласуются полученные нами
В литературе также описано снижение чувстви- результаты: в отсутствие в культуральной системе
тельности эпителия эндометрия к гормонам в эпителия и без моделирования децидуализации
2D культуре (Olalekan et al., 2017). Мы предпола- стромы уровень Esr1 в ней закономерно не изме-
гаем, что полученный результат может быть свя- няется.
зан с необходимостью совместного культивиро- В работе, посвященной изучению зависимо-
вания эпителиальных клеток со стромой, так как сти экспрессии рецепторов к половым гормонам
ответ на гормональную стимуляцию в эндомет- от стадий эстрального цикла, было показано, что
рии in vivo во многом определяется взаимодей- интенсивность иммуногистохимической окраски
ствиями стромального и эпителиального ком- ER была максимальной и в эпителии, и в строме
партментов. во время диэструса и проэструса. Для этих стадий
Р4 ингибировал экспрессию Esr1 в клетках характерно высокое содержание Е2 в сыворотке
эпителия по сравнению с контролем уже на 2 сут крови (Mote et al., 2006). Мы не обнаружили пря-
культивирования (p = 0.028). Этот эффект сохра- мой связи между воздействием Е2 и повышением
нялся и к 5 сут (p = 0.023) (рис. 8). Наши результа- уровня экспрессии Esr1 в эпителии и строме. Од-
ты укладываются в концепцию того, что исходно нако стоит учитывать, что в этой работе уровень
высокий уровень Esr1 в эпителии снижается за экспрессии рецепторов оценивали при помощи
счет гормонального воздействия, обусловленного иммуногистохимических методов, которые не яв-
ОНТОГЕНЕЗ том 52 №6 2021ВЛИЯНИЕ ГОРМОНАЛЬНОГО ФОНА НА ЭКСПРЕССИЮ РЕЦЕПТОРОВ 463
Эпителий Pgr Строма Pgr
0.0003 0.003
0.0002 0.002
rQ
0.0001 0.001
0 0
5 я
E2 ей
5 я
P4 ей
5 я
P ей
5 я
ей
5 я
E2 ей
5 я
P4 ей
5 я
P ей
5 я
ей
Б / 2 дн
E2 дн
P4 дн
P4 дн
Б / 2 дн
E2 дн
P4 дн
P4 дн
дн
дн
E2 дн
дн
дн
дн
E2 дн
дн
2
2
T2 4 2
2
2
T2 4 2
Г
Г
Г
Г
Б/
Б/
Тип гормонального воздействия Тип гормонального воздействия
Рис. 9. Анализ экспрессии гена рецептора к прогестерону в культурах клеток эпителия и стромы, выделенных из эн-
дометрия матки и культивируемых при разных гормональных режимах в течение 2 и 5 сут. Уровень экспрессии нор-
мализован на экспрессию референсного гена ActinB.
ляются оптимальными для точной оценки уровня ванном в культуре и разобщенном с децидуализи-
экспрессии ядерных рецепторов. Измерения про- рующейся стромой ЛЭ не происходило снижение
водили с интервалами 12–24 часа, эстральный уровня экспрессии PR, которое in vivo обусловле-
цикл мыши нерегулярен и длится 4–5 дней, по- но взаимодействием со стромой.
этому для измерения уровня экспрессии рецепто-
ров желательны более короткие интервалы между В работе, посвященной изучению экспрессии
измерениями. рецепторов к половым гормонам на протяжении
стадий эстрального цикла, показали, что в строме
В строме как на 2, так и на 5 сут, уровень экс- интенсивность окраски обеих изоформ PR изме-
прессии Pgr был значительно выше, чем в эпите- нялась в течение эстрального цикла с пиком ин-
лии (p < 0.05). Это ожидаемый результат, так как тенсивности в раннем проэструсе и эструсе, что
строма эндометрия в большей степени является соотвествует максимальной секреции Е2 (Mote
Р4 зависимой тканью, чем эпителий. et al., 2006). В позднем проэструсе интенсивность
И в эпителии, и в строме экспрессия Pgr имела окраски была минимальной, а концентрация P4 в
схожую динамику. На 2 сут уровень экспрессии плазме крови максимальной (Mote et al., 2006).
Pgr не изменялся вне зависимости от режима гор-
монального воздействия. На пятые сутки экс- Несмотря на то, что статистически значимых
прессия Pgr возрастала, как в эпителии (p < 0.01), отличий выявлено не было, мы также обнаружи-
так и в строме (p < 0.01). ли тенденцию к тому, что Р4 в большей степени,
чем другие гормональные режимы, ингибирует
Для мыши было показано, что Pgr экспресси- рост уровня экспрессии PR к 5 сут в обеих культу-
ровался в ЛЭ в околоимплантационные сроки, рах (рис. 9).
уровень его экспрессии снижался с началом де-
цидуализации стромы. При этом сигнальный В другом исследовании подтверждается ре-
путь, индуцируемый Р4 через PR был необходим зультат работы Мота с соавторами, полученный
для прохождения имплантации. На этапе, когда уже при изучении матки мыши в начале беремен-
децидуальная зона была сформирована, экс- ности. В околоимплантационный период фолли-
прессия PR в эпителии уже не детектировалась. кулярный Е2 обеспечивал подготовку клеток стро-
В то же время у псевдобеременных мышей сни- мы к децидуализации и активировал экспрессию
жение уровня экспрессии PR в эпителии прохо- PRs в эндометрии (Zhang et al., 2013). Мы не наблю-
дило только, если искусственно была индуциро- дали такого эффекта. В группе Е2 не происходило
вана децидуальная реакция. Таким образом, пре- роста уровня экспрессии рецепторов на 5 день,
кращение экспрессии PR в эпителии эндометрия превышающего рост в контрольной группе без
регулируется локальными факторами, продуци- добавления гормонов. Этот результат может объ-
руемыми в матке, а также за счет молекулярного ясняться тем, что Е2-зависимый рост PR в клет-
диалога между стромальным и эпителиальным ках стромы in vivo обусловлен взаимодействием с
компартментами (Honglu et al., 2012). Этот факт эпителием. Возможно также в среде не хватало
согласуется с нашими результатами: в изолиро- других про-децидуальных факторов, необходи-
ОНТОГЕНЕЗ том 52 №6 2021464 ИЗМАЙЛОВА и др.
мых для увеличения уровня экспрессии PR в БЛАГОДАРНОСТИ
строме. Работа выполнена с использованием оборудова-
ния центра коллективного пользования (ЦКП) ИБР
им. Н.К. Кольцова РАН.
ЗАКЛЮЧЕНИЕ
Суммируя полученные результаты и проведен- ФИНАНСИРОВАНИЕ РАБОТЫ
ный анализ литературы, можно заключить, что
Работа выполнена за счет гранта Российского На-
контроль экспрессии рецепторов к половым гор- учного Фонда (проект № 21-74-30015).
монам в большинстве случаев не сводится к про-
стой положительной или отрицательной обрат-
ной связи между гормональным сигналом и изме- СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
нением уровня экспрессии его рецептора. Не все
Все применимые международные, национальные
результаты объясняются также и влиянием E2 на и/или институциональные принципы использования
экспрессию PR и Р4 на экспрессию ЕR. Почти животных в экспериментах и условия ухода за ними бы-
точно можно заключить, что молекулярный диа- ли соблюдены. Люди в данном исследовании не участ-
лог между эпителием и стромой эндометрия игра- вовали в качестве объектов.
ет роль в регуляции экспрессии ЕR и PR (Honglu
et al., 2012). Также на это влияют и другие факто-
ры: сигналы, поступающие от эмбриона, дециду- КОНФЛИКТ ИНТЕРЕСОВ
ализация стромы. Показано, что различается ре- Авторы заявляют, что конфликт интересов отсут-
гуляция экспрессии разных изоформ рецепторов ствует.
(Mote et al., 2006).
Решение этого уравнения со множеством не- ИНФОРМАЦИЯ О ВКЛАДЕ АВТОРОВ
известных необходимо для разработки адекват-
ных моделей имплантации и для разработки тера- Любовь Измайлова и Анна Гайдамака выполняли
певтических подходов для заболеваний, ассоции- эксперименты и написали текст статьи, Олеся Серби-
рованных с бесплодием. Затрудняет работу в этом на выделила РНК из всех образцов, Екатерина Вороте-
направлении то, что часть более ранних результа- ляк планировала работу и отредактировала текст.
тов получена с использованием устаревших мето-
дов и может считаться достоверной лишь отчасти СПИСОК ЛИТЕРАТУРЫ
(Tan et al., 1999; Mote et al., 2006). Наши результа-
ты в целом согласуются с полученными ранее. Од- Зурина И.М. Сравнительное исследование эпителио-
мезенхимной пластичности соматических клеток
нако, они ясно показывают необходимость экспе-
человека в условиях 3D культивирования: Автореф.
риментов, в которых гормональное влияние будет дис. ... канд. биол. наук. М.: Ин-т общей патологии
изучаться на системах с сокультивированием эпи- и патофизиологии, 2017. 176 с.
телия и стромы. Также расширить понимание во-
проса может помочь применение методов секвени- Aguilera-Castrejon A., Oldak B., Shani T. et al. Ex utero
mouse embryogenesis from pre-gastrulation to late or-
рования для оценки изменений транскриптома,
ganogenesis // Nature. 2021. V. 593. P. 119–124.
вызванных гормональной стимуляцией. Кроме то-
го, более достоверные данные могут быть получе- Bedzhov I., Leung C.Y., Bialecka M., Zernicka-Goetz M.
ны путем анализа 3D культуральных систем, в ко- In vitro culture of mouse blastocysts beyond the implan-
торых клетки находились бы в более привычных им tation stages // Nat. Protoc. 2014. V. 9. № 12. P. 2732–
2739.
условиях.
Chávez D.J., Anderson T.L. The Glycocalyx of the mouse
Результаты по изучению полярности клеток uterine luminal epithelium during estrus, early pregnan-
эпителия эндометрия также говорят в пользу cy, the peri-implantation period, and delayed implanta-
применения системы 3D культивирования для tion. acquisition of ricinus communis binding sites
более точной оценки влияния гормонов на ТЦМ. during pregnancy // Biol. Reprod. 1985. V. 32. № 1135.
При этом для исследования подвижности клеток P. 42.
возможно применение 2D моделей ввиду выра- Clercq K. De, Hennes A., Vriens J. Isolation of mouse endo-
женного ответа эпителиальных клеток на гормо- metrial epithelial and stromal cells for in vitro decidual-
нальное воздействие. В дополнение к этому, для ization // J. Vis. Exp. 2017. V. 2017. № 121. P. 1–10.
исследования механизмов ТЦМ необходимо ис- Du F., Yang R., Ma H.L. et al. Expression of transcriptional
пользовать нативные клетки эпителия эндомет- repressor SNAI2 gene in mouse endometrium and its ef-
рия, но не опухолевые клетки, для которых свой- fect during embryo implantation // Appl. Biochem.
ственен ЭМП. Biotechnol. 2009. V. 157. P. 346–355.
ОНТОГЕНЕЗ том 52 №6 2021ВЛИЯНИЕ ГОРМОНАЛЬНОГО ФОНА НА ЭКСПРЕССИЮ РЕЦЕПТОРОВ 465
Gou J., Hu T., Li L. et al. Role of epithelial-mesenchymal estrous cycle // Endocrinology. 2006. V. 147. № 12.
transition regulated by twist basic helix-loop-helix tran- P. 5503–5512.
scription factor 2 (Twist2) in embryo implantation in
Nieto M.A., Huang R.Y.J., Jackson R.A. et al. // Cell. 2016.
mice // Reprod. Fertil. Dev. 2019. V. 31. № 5. P. 932–
V. 166. № 1. P. 21–45.
940.
Hamidi H., Lilja J., Ivaska J. Using xCELLigence RTCA Olalekan S.A., Burdette J.E., Getsios S. et al. Development
Instrument to Measure Cell Adhesion // Bio-Protocol. of a novel human recellularized endometrium that re-
2017. V. 7. № 24. P. 1–17 sponds to a 28-day hormone treatment // Biol. Reprod.
Hantak A.M., Bagchi I.C., Bagchi M.K. Role of uterine stro- 2017. V. 96. № 5. P. 971–981.
mal-epithelial crosstalk in embryo implantation // Int. Quinn C.E., Matson B.C., Wetendorf M. et al. Pinopodes:
J. Dev. Biol. 2014. V. 58. № 2–4. P. 139–146. Recent advancements, current perspectives, and future
Honglu D., Paria B.C., Xiao S. et al. Temporal expression directions // Mol. Cell Endocrinol. 2019. V. 5. P. 501.
pattern of progesterone receptor in the uterine luminal P. 110644.
epithelium suggests its requirement during early events
of implantation // Bone. 2012. V. 23. № 1. P. 1–7. Shakya S., Asosingh K., Mack J.A. et al. Optimized protocol
for the preparation of single cells from cutaneous
Illingworth I.M., Kiszka I., Bagley S. et al. Desmosomes are wounds for flow cytometric cell sorting and analysis of
reduced in the mouse uterine luminal epithelium during macrophages // Methods. X. 2020. V. 7. P. 101027.
the preimplantation period of pregnancy: a mechanism
for facilitation of implantation // Biol. Reprod. 2000. Tan J., Paria B.C., Dey S.K. et al. Differential uterine ex-
V. 63. P. 1764–1773. pression of estrogen and progesterone receptors cor-
Kallen A.N., Haines K., Taylor H.S. HOXA10 regulates ex- relates with uterine preparation for implantation and de-
pression of cytokeratin 15 in endometrial epithelial cyto- cidualization in the mouse // Endocrinology. 1999.
skeletal remodeling // Reprod. Sci. 2014. V. 21. № 7. V. 140. № 11. P. 5310–5321.
P. 892–897. Tian H., Xu J.Y., Tian Y. et al. A cell culture condition that
Kaneko Y., Day M.L., Murphy C.R. Integrin β3 in rat blasto- induces the mesenchymal-epithelial transition of dedif-
cysts and epithelial cells is essential for implantation ferentiated porcine retinal pigment epithelial cells // Ex-
in vitro: Studies with Ishikawa cells and small interfering perimental Eye Research. 2018. V. 177. P. 160–172.
RNA transfection // Hum. Reprod. 2011. V. 26. № 7.
P. 1665–1674. Uchida H., Maruyama T., Oda H. Studies using an in vitro
model show evidence of involvement of epithelial-mes-
Kaneko Y., Lecce L., Day M.L. et al. B1 and B3 integrins dis- enchymal transition of human endometrial epithelial
assemble from basal focal adhesions and b3 integrin is cells in human embryo implantation // J. Biol. Chem.
later localised to the apical plasma membrane of rat 2012. V. 287. P. 4441–4450.
uterine luminal epithelial cells at the time of implanta-
tion // Reprod. Fertil. Dev. 2011. V. 23. № 3. P. 481– Vasquez Y.M., DeMayo F.J. Role of nuclear receptors in
495. blastocyst implantation // Semin. Cell Dev. Biol. 2013.
Kubaczka C., Senner C., Araúzo-Bravo M.J. et al. Derivation V. 24. № 10–12. P. 724–735.
and maintenance of murine trophoblast stem cells under Wang H., Dey S.K. Roadmap to embryo implantation:
defined conditions // Stem Cell Reports. 2014. V. 2. Clues from mouse models // Nat. Rev. Genet. 2006.
№ 2. P. 232–242. V. 7. № 3. P. 185–199.
Lamouille S., Xu J., Derynck R. Molecular mechanisms of
epithelial-mesenchymal transition // Nat. Rev. Mol. Whitby S., Zhou W., Dimitriadis E. Alterations in epithelial
Cell Biol. 2014. V. 15. № 3. P. 178–196. cell polarity during endometrial receptivity: A systemat-
ic review // Front. Endocrinol. (Lausanne). 2020. V. 11.
Lin P. F., Lan X.Li., Chen F.L. et al. Reference gene selec- P. 1–14.
tion for real-time quantitative PCR analysis of the
mouse uterus in the peri-implantation period // PLoS Wilcox A.J., Weinberg C.R., O’Connor J.F. et al. Incidence of
One. 2013. V. 8. № 4. P. 4–11. early loss of pregnancy // N. Engl. J. Med. 1988. V. 319.
Liu, Z., Qi, S., Zhao, X. et al. Metformin inhibits 17β-estra- № 4. P. 189–194.
diol-induced epithelial-to-mesenchymal transition via Ye T.M., Pang R.T.K., Leung C.O.N. et al. Development and
βKlotho-related ERK1/2 signaling and AMPKα signal- characterization of an endometrial tissue culture model
ing in endometrial adenocarcinoma cells // Oncotarget. for study of early implantation events // Fertil. Steril.
2016. V. 7. № 16. P. 21315–21331. 2012. V. 98. № 6. P. 1581–1589.
Luddi A., Pavone V., Semplici B. et al. Organoids of human
endometrium: A powerful in vitro model for the endo- Ye X. Uterine luminal epithelium as the transient gateway
metrium-embryo cross-talk at the implantation site // for embryo implantation // Trends Endocrinol. Metab.
Cells. 2020. V. 9. № 5. P. 1121. 2020. V. 31. № 2. P. 165–180.
Mote P.A., Arnett-Mansfield R.L., Gava N. et al. Overlap- Zhang S., Lin H., Kong S. et al. Physiological and molecular
ping and distinct expression of progesterone receptors A determinants of embryo implantation // Mol. Aspects
and B in mouse uterus and mammary gland during the Med. 2013. V. 34. № 5. P. 939–980.
ОНТОГЕНЕЗ том 52 №6 2021466 ИЗМАЙЛОВА и др.
Influence of Sex Steroids on the Expression of Its Receptors and Migratory Potential
of Mouse Epithelial and Stromal Endometrial Cells In Vitro
L. Sh. Izmailova1, *, А. A. Gaidamaka1, О. О. Serbina1, and E. A. Vorotelyak1
1
Koltzov Institute of Developmental Biology of the Russian Academy of Sciences, ul. Vavilova 26, Moscow, 119334 Russia
*e-mail: luba.ranaway-94@yandex.ru
Preparation for embryo implantation is a complex process that includes changes at both cellular and tissue
levels in the epithelium and stroma of the endometrium. The success of future pregnancy depends on the reg-
ulation of preimplantation changes in the endometrium and during the onset of the receptive period. Model-
ing the regulation of endometrial functioning in vitro makes it possible to study the process of preparing the
endometrium for embryo implantation, which is especially important for the development of therapeutic ap-
proaches for the treatment of diseases associated with infertility. In this work we show the effects of sex ste-
roids on the changes in endometrial epithelium polarity in vitro. Changes in endometrial epithelium polarity
are also an important process during in vivo implantation. The mutual influence of estrogen and progester-
one, the main regulators of endometrium, on the expression of its receptors is also described.
Keywords: endometrium, epithelium, stroma, epithelial-mesenchymal transition, sex steroids
ОНТОГЕНЕЗ том 52 №6 2021Вы также можете почитать