ФЕНОТИПИЧЕСКОЕ РАЗНООБРАЗИЕ И ФУНКЦИОНАЛЬНЫЙ ПОТЕНЦИАЛ МЕТАНО- И МЕТИЛОТРОФНЫХ БАКТЕРИЙ СЕМЕЙСТВА BEIJERINCKIACEAE
←
→
Транскрипция содержимого страницы
Если ваш браузер не отображает страницу правильно, пожалуйста, читайте содержимое страницы ниже
На правах рукописи
Воробьев Алексей Викторович
ФЕНОТИПИЧЕСКОЕ РАЗНООБРАЗИЕ И ФУНКЦИОНАЛЬНЫЙ
ПОТЕНЦИАЛ МЕТАНО- И МЕТИЛОТРОФНЫХ БАКТЕРИЙ
СЕМЕЙСТВА BEIJERINCKIACEAE
Специальность 03.00.07 – микробиология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Москва - 2009
Работа выполнена в Учреждении Российской академии наук Институт
микробиологии им. С.Н. Виноградского РАН
Научный руководитель: доктор биологических наук
С.Н. Дедыш
Официальные оппоненты: доктор биологических наук
Д.Ю. Сорокин
доктор биологических наук
А.Л. Степанов
Ведущая организация: Учреждение Российской академии наук
Институт биохимии и физиологии
микроорганизмов им. Г.К. Скрябина РАН
Защита диссертации состоится «30» ноября 2009 г. в 1200 ч. на заседании
диссертационного совета Д.002.224.01 в Учреждении Российской академии
наук Институт микробиологии им. С.Н. Виноградского РАН по адресу: 117312,
г. Москва, Проспект 60-летия Октября, д. 7, к. 2.
С диссертацией можно ознакомиться в библиотеке Учреждения Российской
академии наук Институт микробиологии им. С.Н. Виноградского РАН.
Автореферат разослан «___» октября 2009 г.
Ученый секретарь
диссертационного совета,
кандидат биологических наук Т.В. Хижняк
Актуальность проблемы.
Аэробные метано- и метилотрофные бактерии – это уникальные группы
прокариотных микроорганизмов, способных использовать метан и его
одноуглеродные производные в качестве единственного источника углерода и
энергии (Söhngen, 1906; Anthony, 1982; Whittenbury, Krieg, 1984; Hanson, Hanson,
1996; Bowman, 2000; Гальченко, 2001; Lidstrom, 2006). Ключевым ферментом
метанотрофов является метанмонооксигеназа (ММО), катализирующая окисление
СН4 до CH3OH и существующая в двух формах – мембранной (мММО) и
растворимой (рММО). Окисление метанола до формальдегида осуществляется
другим ключевым ферментом – метанолдегидрогеназой (МДГ), которым обладают
как метанотрофы, так и метилотрофные бактерии. Эти микроорганизмы формируют
естественный бактериальный фильтр, регулирующий поступление метана в
атмосферу, а также определяют баланс С1-соединений в природных экосистемах
(Заварзин, 1979, 1984; Conrad, 1996).
Метилотрофия широко распространена среди представителей различных
филогенетических групп домена Bacteria, тогда как способность к росту на метане на
настоящий момент выявлена лишь у организмов двух групп, Proteobacteria и
Verrucomicrobia (Hanson, Hanson, 1996; Op den Camp et al., 2009). Метанотрофные
Verrucomicrobia открыты недавно, и сведения об их биологии пока ограничены.
Гораздо лучше изучены метанотрофные представители Proteobacteria, формирующие
филогенетически тесные кластеры в пределах Alpha- и Gammaproteobacteria и не
обнаруживающие близкого родства с гетеротрофными бактериями. Единственным
исключением является семейство Beijerinckiaceae класса Alphaproteobacteria, которое
объединяет умеренно ацидофильные, аэробные бактерии с различными типами
метаболизма – гетеротрофов, метано- и метилотрофов. С момента формирования
(Alston, 1936; Starkey et al., 1939) и до недавнего времени все представители этого
семейства считались гетеротрофными азотфиксаторами, использующими широкий
спектр органических соединений. Эти представления изменились после описания
умеренно ацидофильных метанотрофных бактерий родов Methylocella и Methylocapsa,
которые оказались ближайшими филогенетическими родственниками гетеротрофов
рода Beijerinckia (Dedysh et al., 2000, 2002). Единственный описанный представитель
рода Methylocapsa, Methylocapsa acidiphila, является облигатным метанотрофом,
обладающим мММО. Род Methylocella ныне насчитывает три вида, M. palustris, M.
silvestris и M. tundrae (Dedysh et al., 2000, 2004, Dunfield et al., 2003), представители
которых, в отличие от остальных ныне известных метанотрофов, обладают одной
только рММО. Еще одной уникальной для метанотрофов особенностью бактерий
рода Methylocella является их способность к росту не только на С1-соединениях, но и
на ацетате, пирувате, сукцинате, малате и этаноле (Dedysh et al., 2005а; Theisen et al.,
2005). Представления о метаболическом потенциале бактерий рода Beijerinckia также
были пересмотрены после выявления у B. mobilis способности к автотрофной
1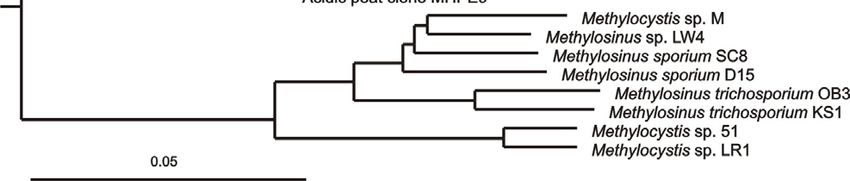
метилотрофии (Dedysh et al., 2005б). Итак, в настоящий момент семейство
Beijerinckiaceae включает в себя истинных гетеротрофов, облигатных метанотрофов,
а также организмы с различными типами “промежуточного” метаболизма –
факультативных метано- и метилотрофов.
Применение молекулярных подходов для изучения экологии микроорганизмов
выявило чрезвычайно широкое распространение представителей Beijerinckiaceae. Это
объясняется тем, что большинство наземных экосистем Северного полушария
характеризуются кислой реакцией среды, к которой хорошо адаптированы организмы
этого семейства. Нуклеотидные последовательности генов 16S рРНК представителей
Beijerinckiaceae были обнаружены в лесных почвах и почвах агроценозов, в болотах
различных типов, а также в покрывающих почвах свалок (Radajewski et al., 2000,
2002; Morris et al., 2002; Knief et al., 2003; Raghoebarsing et al., 2005; Lau et al., 2007;
Cebron et al., 2007; Chen et al., 2007, 2008). Анализ этих последовательностей, однако,
не позволяет сделать однозначных заключений о метаболическом типе тех
организмов, которым они принадлежат, из-за гетерогенности этой группы бактерий.
Количество культивируемых метилотрофных представителей этого семейства также
очень мало, что объясняется, в частности, сложностью работы по их выделению.
Таким образом, микробные агенты окисления С1-соединений в кислых наземных
экосистемах остаются изучены недостаточно. Устранению этого пробела было
посвящено настоящее исследование, направленное на расширение спектра
культивируемых метано- и метилотрофных представителей семейства Beijerinckiaceae
и изучению их метаболического потенциала и особенностей биологии.
Цели и задачи исследования.
Цель работы – изучение фенотипического и филогенетического разнообразия
метано- и метилотрофных бактерий семейства Beijerinckiaceae и выявление различий
в метаболическом потенциале и экологических стратегиях бактерий этой группы.
Для достижения этой цели нами были поставлены следующие задачи:
1. Получение новых изолятов метано- и метилотрофных бактерий семейства
Beijerinckiaceae, изучение их физиологии, спектра используемых субстратов, а
также составление полной таксономической характеристики этих бактерий.
2. Определение функционального потенциала и ростовых характеристик
облигатно- и факультативно- метанотрофных представителей семейства
Beijerinckiaceae для выявления различий экологических стратегий,
используемых этими организмами.
3. Оценка роли факультативных метанотрофов в общей структуре бактериального
метанокисляющего фильтра болотных экосистем.
Научная новизна и значимость работы.
Значительно расширено разнообразие культивируемых представителей
семейства Beijerinckiaceae с метано- и метилотрофными типами метаболизма.
2
Показано, что эти бактерии широко распространены в наземных экосистемах с
кислой реакцией среды. Впервые выявлен существенный вклад факультативных
метанотрофов в общую метанокисляющую активность торфа сфагновых болот.
Описан и узаконен новый род и вид облигатно ацидофильных метилотрофных
бактерий семейства Beijerinckiaceae - Methylovirgula ligni gen. nov., sp. nov. Два новых
таксона факультативно метанотрофных бактерий семейства Beijerinckiaceae -
‘Methyloferula sphagni’ gen. nov., sp. nov. и ‘Methylocapsa aurea’ sp. nov. находятся в
процессе узаконивания. Типовые штаммы этих бактерий депонированы в
международных коллекциях микроорганизмов DSMZ, NCIMB, LMG и ВКМ.
Практическая значимость.
Показано широкое распространение факультативной метанотрофии в микробном
мире, что вносит коррективы в ранее принятое представление о том, что доступность
СН4 является единственным энергетическим фактором, регулирующим процессы его
микробного окисления в природе.
Существенно дополнена база данных нуклеотидных последовательностей генов
16S рРНК, pmoA, mmoX, mxaF и nifH метилотрофных бактерий, которая может быть
использована для разработки молекулярных методов детекции этих
микроорганизмов, основанных на использовании ПЦР или микрочипов.
Получены новые культуры ацидофильных метанотрофных микроорганизмов,
обладающих растворимой формой ММО, имеющих хороший потенциал для
применения в биотехнологии очистки кислых природных сред от загрязнения
галогенированными алканами и ароматическими углеводородами.
Апробация работы.
Материалы диссертации доложены и обсуждены на международных и
российских конференциях и симпозиумах:
1. Международной молодежной школе-конференции «Актуальные аспекты
современной микробиологии: III Международная молодежная школа-
конференция», Москва, ИНМИ РАН, 2007.
2. 12th International Symposium on Microbial Ecology, Cairns, Australia. August 17-22,
2008.
3. XVI международной конференции студентов, аспирантов и молодых ученых
«Ломоносов», Москва, МГУ, 2009.
4. 3th Congress of European microbiologists. Gothenburg, Sweden, June 28 – July 2, 2009.
Публикации.
Материалы диссертации содержатся в 7 печатных работах: 2 экспериментальных
статьях и 5 тезисах.
Объем и структура диссертации.
3
Диссертация состоит из введения, глав, заключения и выводов, изложенных на
_131_страницах, включая _9_ таблиц, _37_ рисунков и списка литературы из _176_
наименований, из них _12_ – на русском и _164_ – на английском языке.
Место проведения работы и благодарности.
Работа была выполнена в лаборатории Микробиологии болотных экосистем
отдела Микробных сообществ Учреждения Российской академии наук Институт
микробиологии им. С.Н. Виноградского РАН с 2006 по 2009 годы под
руководством д.б.н. С.Н. Дедыш.
Образцы торфа болот различного географического расположения были
предоставлены автору к.б.н. И.С. Куличевской, к.б.н. Т.А. Панкратовым (ИНМИ
РАН), а также отобраны самим автором.
Исследованные в работе изоляты метано- и метилотрофных бактерий были
получены и предоставлены автору д.б.н. С.Н. Дедыш (ИНМИ РАН), P. Dunfield
(University of Calgary, Canada), L. Folman (NIOO-KNAW, The Netherlands), а также
были выделены самим автором. Исследования ультратонкого строения клеток
метилотрофов были проведены совместно с к.б.н. Н.И. Сузиной (ИБФМ РАН, г.
Пущино). Анализ ферментного состава метилотрофных бактерий был проведен д.б.н.
Дорониной Н.В. (ИБФМ РАН, г. Пущино). Анализ хинонов был проведен к.х.н. Б.П.
Баскуновым (ИБФМ РАН, г. Пущино). ДНК-ДНК гибридизация и определение
содержания Г+Ц пар в ДНК проведены к.б.н. Е.Н. Детковой (ИНМИ РАН).
Работа была выполнена при финансировании в рамках программ Президиума
РАН «Молекулярная и клеточная биология» и «Биоразнообразие», проекта РФФИ-
ННИО 09-04-91332, а также работ по Гос. контракту с Роснаукой № 02.740.11.0023.
Автор выражает искреннюю благодарность всем вышеупомянутым участникам
данной работы, а также особую признательность д.б.н. С.Н. Дедыш и акад. Г.А.
Заварзину за полезные практические советы и поддержку на всех этапах работы.
СОДЕРЖАНИЕ РАБОТЫ
Материалы и методы исследования
Образцы торфа для получения накопительных культур метано- и
метилотрофов, а также для количественного учета клеток этих микроорганизмов
были отобраны из сфагновых болот Торфяное, Архангельской обл. (65º01'N, 35º44'E),
и Бакчарское, Томской обл. (56º51’N, 82º51’E).
Накопительные культуры метанотрофных бактерий получали путем
инокулирования торфом жидкой минеральной среды М2 (pH 5.0-5.5) следующего
состава (г/л): MgSO4×7H2O-0.05; KH2PO4-0.1; CaCl2×2H2O-0.01; NaCl-0.02; раствор
микроэлементов для метанотрофов (Гальченко, 2001) - 1мл. В качестве источника
4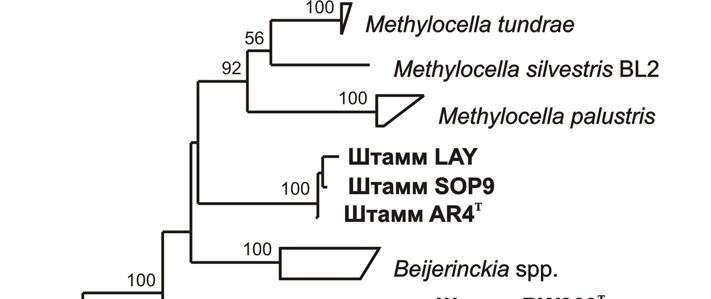
азота в среду вносили (г/л): (NH4)2SO4-0.05, KNO3-0.1 или же использовали среду М2
без источника связанного азота. Для культивирования метилотрофных бактерий в
среду добавляли метанол в концентрациях 0.01-5 об. %, тогда как для метанотрофов в
газовую фазу опытных флаконов вводили метан до 10-20 об. % и инкубировали их в
статических условиях или на шейкере (120 об./мин.). Чистые культуры метано- и
метилотрофных бактерий выделяли путем многократных рассевов на плотной среде
М2 с высокоочищенным агар-агаром, Gellun Gum или PhytaGel, или методом
предельных разведений в жидких средах вышеуказанного состава.
Объектами настоящего исследования являлись 8 штаммов метано- и
метилотрофных бактерий. Штаммы KYG и LAY были предоставлены P. Dunfield
(University of Calgary, Canada), штаммы BW863 и BW872 - L. Folman (NIOO-KNAW,
The Netherlands), штамм AR4 - Дедыш С.Н. (ИНМИ РАН). Штаммы SOP9, RP01 и
ESY были выделены автором работы.
В качестве тест-организмов использовали культуры Beijerinckia indica (ATCC
9039 ), Beijerinckia mobilis (DSM 2326Т), Methylocapsa acidiphila (DSM 13967Т),
Т
Methylocella palustris (ATCC 700799Т), Methylocella silvestris (DSM 15510Т) и
Methylocella tundrae (DSM 15673 Т).
Культивирование изолятов проводили на вышеописанной среде М2, а также
на пятикратно разведенной среде NMS (Dunfield et al., 2003). Динамику роста
культур прослеживали путем определения оптической плотности суспензий на
Eppendorf Biophotometer (длина волны 600 нм). Для определения ростового диапазона
и оптимума pH изолятов использовали среды, варьирование кислотности которых
осуществляли добавлением 0.1М растворов H2SO4 и NaOH. Активность окисления
метана образцами торфа и чистыми культурами метанотрофных бактерий определяли
газохроматографически на хроматографе Кристалл 5000 (“ЗАО Хроматэк”, Россия) с
пламенно-ионизационным детектором.
Выделение ДНК из клеток микроорганизмов производили по ранее описанной
методике (Dedysh et al., 1998). В ходе работы производили ПЦР-амплификацию
фрагментов следующих филогенетических и функциональных генов метано- и
метилотрофов: а) гена 16S рРНК изолятов с использованием универсальных
праймеров, предложенных Weisburg et al. (1991); б) гена pmoA, кодирующего
полипептид, несущий активный центр мембраной ММО (Holmes et al., 1995; Kolb et
al., 2005); в) гена mmoX, кодирующего α-субчастицу растворимой ММО (Auman et al.,
2000); г) гена mxaF, кодирующего большую субъединицу МДГ (McDonald, Murrell,
1997); д) гена nifH, кодирующего редуктазный компонент нитрогеназного комплекса
(Auman et al., 2001; Булыгина и др., 2002). ПЦР реакции проводили на термоциклере
PE GeneAmp PCR System 9700 (“Perkin-Elmer Applied Biosystems”, США). Проверку
продуктов ПЦР осуществляли путем их электрофореза в 1.1% агарозном геле с
последующим окрашиванием бромистым этидием и визуализацией продуктов
реакции с помощью УФ-трансиллюминатора.
5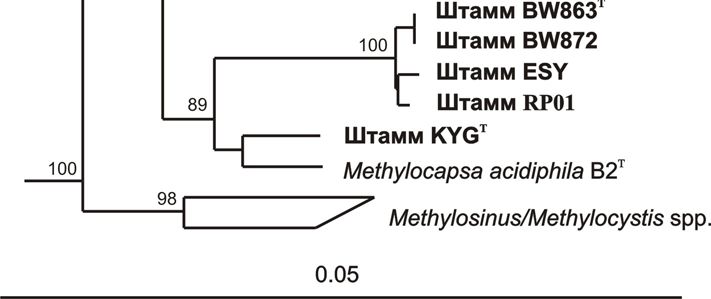
Редактирование полученных нуклеотидных последовательностей
осуществляли с использованием программы SeqMan (Lasergene 7.0; DNA Star
Package). Сравнение полученных последовательностей с таковыми в базе данных
GenBank проводили с использованием программы Blast2 (http://www.ebi.ac.uk/blast2/).
Построение филогенетических дендрограмм производили с использованием
программного пакета ARB (http://www.arb-home.de). Статистическую достоверность
дендрограмм рассчитывали с использованием программного пакета Phylip с помощью
“bootstrap”-анализа путем построения 1000 альтернативных деревьев.
Поиск уникальных последовательностей в 16S рРНК новых изолятов
метанотрофных бактерий и создание олигонуклеотидных зондов для их детекции
осуществляли с использованием функции “Probe Design” пакета ARB.
Специфичность зондов проверяли путем тестирования в базе данных Ribosomal
Database Project (http://rdp.cme.msu.edu). Синтез зондов, меченых флуоресцентным
красителем Cy3, осуществлялся компанией “Синтол” (Москва, Россия).
Экстракцию микробных клеток из сфагнового торфа для дальнейшего анализа
методом FISH проводили с использованием гомогенизатора BagMixer 100 “MiniMix”
(Interscience, Франция) и прилагающихся к нему стерильных пакетов BagFilter®. К 2 г
торфа добавляли 20 мл стерильной дистиллированной воды, обрабатывали с
помощью гомогенизатора в течение 10 минут при максимальной частоте. Затем
отбирали 0.5 мл полученной суспензии и фиксировали с использованием 4%-го
раствора формальдегида в фосфатном буфере (NaCl – 8.0 г, KCl – 0.2 г, Na2HPO4 –
1.44 г, NaH2PO4 – 0.2 г, H2O – 1 л, pH 7.0) в течение 1.5 часов. Образец осаждали и
промывали фосфатным буфером. Фиксированные образцы ресуспендировали в
растворе 100% этанола и фосфатного буфера (1:1, об:об) и до анализа хранили при
20ºС.
Для гибридизации использовали набор 16S рРНК-специфичных
олигонуклеотидных зондов: зонды для детекции метанотрофов I и II типов (Eller et
al., 2001), зонды для дифференцированной детекции метанотрофов группы
Methylocystis-Methylosinus, а также бактерий родов Methylocapsa и Methylocella
(Дедыш, 2006). Гибридизацию препаратов с зондами проводили в соответствии с
методикой Stahl & Amann (1991) при температуре 46ºС или 50ºС. Общую численность
микробных клеток в торфе определяли окрашиванием препаратов 1 мкМ раствором
ДНК-специфичного красителя ДАФИ (4’,6’-диамидино-2-фенилиндол).
Препараты анализировали с использованием эпифлуоресцентного микроскопа
Zeiss Axioplan 2 (Йена, Германия) со светофильтрами Zeiss 20 для Cy3-меченных
зондов и Zeiss 02 для окраски ДАФИ. Определение численности целевых групп в
образцах торфа и почвы осуществляли путем учета количества гибридизованных с
зондами клеток в 100 полях зрения, с последующим расчётом численности популяций
на 1 г влажного торфа.
6Результаты исследований
1. Новые изоляты облигатно ацидофильных метилотрофных бактерий
семейства Beijerinckiaceae.
1.1. Изоляты, выделенные из кислой лесной почвы.
В ходе изучения микробного сообщества, участвующего в разложении
древесины бука в кислой (pH 3.3-3.6) лесной почве, L. Folman, сотрудницей
лаборатории W. de Boer (NIOO-KNAW, The Netherlands) было выделено 2 штамма
ацидофильных бактерий – BW863T и BW872 (Folman et al., 2008). Анализ
последовательностей гена 16S рРНК изолятов выявил их принадлежность к семейству
Beijerinckiaceae, причем ближайшим филогенетическим родственником штаммов
BW863T и BW872 из числа ранее описанных организмов была ацидофильная
облигатно метанотрофная бактерия Methylocapsa acidiphila B2 (96.5-97% сходства
последовательностей гена 16S рРНК) (Рис. 1). Так как для дальнейшей
характеристики новых изолятов требовалось их детальное сравнение с M. acidiphila
B2, культуры новых штаммов было переданы для изучения в лабораторию
микробиологии болотных экосистем ИНМИ им. С.Н. Виноградского РАН, где была
выделена и описана вышеупомянутая метанотрофная бактерия.
Рис. 1. Филогенетическая дендрограмма, построенная на основе сравнительного
анализа нуклеотидных последовательностей генов 16S рРНК представителей семейств
Beijerinckiaceae и Methylocystaceae. В качестве внешней группы использованы
нуклеотидные последовательности генов 16S рРНК метанотрофов I типа. Маркер – 0.05
замещений на нуклеотидную позицию. Жирным шрифтом показаны штаммы,
исследованные в настоящей работе.
7Эксперименты по определению спектра утилизируемых субстратов показали,
что, несмотря на близкое родство с Methylocapsa acidiphila B2, штаммы BW863T и
BW872 не были способны к использованию метана, но хорошо росли на метаноле в
диапазоне концентраций 0.01-2 об. % с оптимумом 0.5-1 об. %.
Анализ гена mxaF, кодирующего большую субъединицу МДГ, выявил, что
последовательности MxaF новых штаммов существенно отличались от всех
описанных ранее последовательностей MxaF культивируемых метилотрофов (Рис. 2).
Однако они обнаруживали высокое сходство (95.6-97.5%) с транслированными
аминокислотными последовательностями генов mxaF, выявленными методом
стабильных изотопов (SIP) в кислых лесных почвах, обогащенных 13CH3OH
(Radajewski et al., 2000, 2002) и принадлежащими ранее некультивируемой группе
ацидофильных метилотрофов.
Рис. 2. Дендрограмма, построенная на основе сравнительного анализа
транслированных аминокислотных последовательностей гена mxaF, показывающая
филогенетическое положение штаммов BW863T и BW872 относительно других
метилотрофных представителей Proteobacteria и последовательностей MxaF,
выявленных в кислых почвах методом стабильных изотопов. Маркер, 0.1 замещений
на нуклеотидную позицию.
8Штаммы BW863T и BW872 были представлены грамотрицательными клетками
палочковидной формы, размножающимися бинарным делением. Размер клеток
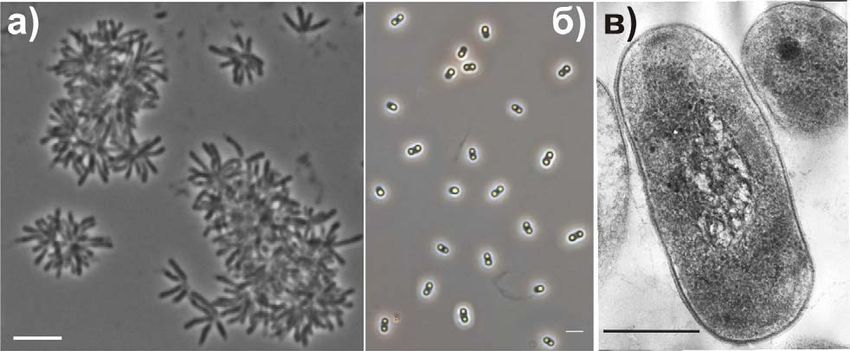
составлял 1.2-2.5 мкм в длину и 0.4-0.65 мкм в ширину (Рис. 3). Клетки, выращенные
на плотной среде, имели тонкие пили-подобные структуры диаметром 5 нм (Рис. 3б.).
Рис. 3. (a) Фазово-контрастная фотография клеток штамма BW863T в 10-суточной
жидкой культуре на среде с метанолом. (б-г) Электронные микрофотографии
ультратонких срезов клеток, выращенных на агаризованной среде. ПГБ, поли-β-
гидроксибутират; П, пили-подобные структуры. Маркеры – 5 мкм (a); 0.5 мкм (б-г).
Кроме метанола, эти изоляты хорошо росли на этаноле (Рис. 4). Слабый рост
наблюдался также на малате, сукцинате и пирувате, однако штаммы не росли на
ацетате, сахарах и других полиуглеродных соединениях. Штаммы BW863T и BW872
являлись мезофильными и облигатно ацидофильными организмами, способными к
росту при pH 3.1-6.5 и в диапазоне температур от +4 до +30˚С, причем оптимальный
рост наблюдался при pH 4.5-5.0 и в интервале температур 15-18ºС (Рис. 5 А, Б).
В составе клеточных жирных кислот у штаммов BW863T и BW872
доминировали 18:1ω7c кислоты, что характерно для представителей родов
Methylocella и Methylocapsa (Табл. 1).
Анализ морфологических, хемотаксономических и генотипических
характеристик новых изолятов позволил установить их принадлежность к новому
роду и виду семейства Beijerinckiaceae - Methylovirgula ligni gen. nov., sp. nov
(Vorob’ev et al., 2009).
9OD600
0.5
0.4 ацетат
малат
пируват
0.3
сукцинат
этанол
0.2 метанол
контроль
0.1
0
0 5 10 15 Сутки
Рис. 4. Рост штамма BW863T на средах с разными источниками углерода.
µ, hr-1 А µ, hr-1 Б
0.05 0.01
0.04 0.008
0.03 0.006
0.02 0.004
0.01 0.002
0 0
2.5 3.5 4.5 5.5 6.5 pH 0 10 20 30 TºC
Рис. 5. Зависимость удельной скорости роста штамма BW863т от величины pH (А) и
температуры (Б) при росте на среде с метанолом.
Характеристика Methylovirgula gen. nov. (Me.thy.lo.vir'gu.la. N.Gr. n. methyl от
Gr. n. methu, вино и Gr. n. hulê, дерево, метильная группа; L. fem. n. virgula, маленькая
палочка; N.L. fem. n. Methylovirgula, метилотрофная палочковидная клетка).
Одиночные, образующие розетки или скопления неправильной формы
грамотрицательные палочки. Размножаются бинарным делением. Колонии выпуклые,
непрозрачные, кремово-белые, диаметром 0.5-1 мм, плотной консистенции с ровным
краем и гладкой поверхностью. Рост в жидких средах гомогенный. Аэробные,
ограниченно факультативные метилотрофы. Предпочтительный субстрат – метанол.
Наблюдается рост на этаноле, а также слабый рост на малате, пирувате, и сукцинате.
Облигатные ацидофилы и мезофилы. Чувствительны к концентрациям NaCl выше
0.7%. Основной хинон - менахинон Q-10. Доминирующая жирная кислота -18:1ω7c.
Содержание пар Г+Ц в ДНК 61.8-62.8 моль %. Род принадлежит к классу
Alphaproteobacteria, семейству Beijerinckiaceae. Типовой вид – Methylovirgula ligni.
10Таблица 1. Состав клеточных жирных кислот новых метило- и метанотрофных
бактерий, выделенных из сфагнового торфа и кислой лесной почвы, а также ранее
описанных представителей родов Methylocella и Methylocapsa.
% общего содержания жирных кислот
Метилотрофы Метанотрофы
Кислота
Methylovirgula ‘Methyloferula Methylocapsa Methylocella spp.**
ligni sphagni’ (AR4T, spp. (M. (M. palustris, M.
(BW863T и SOP9)*** acidiphila B2*, silvestris, M. tundrae)
BW872) *** KYG***)
14:0 0 0.23-0.28 0 0 - 4.1
i15:0 0 0.73-0.85 0.1-0.3 0.2 -1.2
16:0 1.80-1.96 7.89-5.05 5.9-7.3 3.0 - 7.7
16:1ω7c 0.33-0.36 1.82 4.7-6.3 4.7 - 11.3
16:1ω7t 0 0 0 0-5.8
a17:0 0-0.21 0 0 0 – 0.3
17:0 0.53-0.64 0 0.1 0-0.1
i17:0 0-0.23 0.66-0.87 0.6-0.9 0 - 2.5
br17:0 0.23-0.25 0 0 0
cy17:0 0-0.18 1.85 0 0 - 6.5
17:1ω7c 0.16-0.19 0.28 1.0 0
i18:0 0 0.43 0 0 – 0.5
18:0 0.831-0.9 0.61 0.8-7.6 0.4 - 1.2
18:1ω9c 0-0.41 0.32-0.42 0-0.4 0
18:1ω8c 0 0 0 0
18:1ω7c 87.07-92.76 83.22-85.69 78.3-81.5 59.2-82.2
cy19:0 1.39-2.57 0 0 0
cy19:1ω8c 0 1.32-3.77 0 0 - 13.6
20:0 0.11-0.13 0.58 0 0
* Данные Dedysh et al. (2002).
** Данные Dedysh et al. (2000, 2004a) и Dunfield et al. (2003).
*** Определено в настоящем исследовании.
Methylovirgula ligni sp. nov. Methylovirgula ligni (lig'ni. L. gen. n. ligni, из дерева,
изолирован из древесины). Основные характеристики указаны при описании рода.
Размер клеток 1.2-2.5 мкм в длину и 0.4-0.65 мкм в ширину. Источники азота -
аммоний, нитрат, дрожжевой экстракт и мочевина. Нуждаются в дрожжевом
экстракте в качестве фактора роста (50 мг/л среды). Растут при температуре от 4 до
1130˚C, с оптимумом при 15-18˚C, и в диапазоне pH 3.1-6.5, с оптимумом при 4.5-5.0.
Местообитания – болота и почвы с низкими значениями pH. Типовой штамм BW863т
(=DSM 19998T = NCIMB 14408T), изолирован из разлагающейся древесины бука,
инкубированной на поверхности кислой лесной почвы.
1.2 Изоляты, выделенные из сфагновых болот.
В ходе дальнейшей работы из кислых сфагновых болот нами было выделено 2
штамма метилотрофных бактерий, RP01 и ESY, гены 16S рРНК которых
обнаруживали 98.5-99% сходства с таковыми у штаммов BW863T и BW872 (Рис. 1).
Штаммы RP01 и ESY были представлены грамотрицательными клетками
палочковидной формы, размножающимися бинарным делением. Размер клеток
штамма RP01 достигал 1.25-2.6 мкм в длину и 0.4-0.7 мкм в ширину, то есть был
сходным с таковым у штаммов BW863T и BW872. Размеры клеток штамма ESY были
значительно меньше и варьировали в зависимости от условий культивирования. При
росте в оптимальных условиях размеры клеток составляли 0.25-0.6×0.4-2.1 мкм, а
средний объем клеток был равен 0.15 мкм3. При росте в неоптимальных условиях или
на субстратах, не являющихся предпочтительными, клетки штамма ESY были короче
(0.4-1.5 мкм) (Рис. 6), а их объем варьировал в диапазоне 0.02-0.09 мкм3, что
позволяет отнести их к категории ультрамикроформ (Morita, 1981, 1985, 1997).
Рис. 6. Электронно-сканирующая микрофотография клеток штамма ESY. Маркеры – 5
мкм.
Штаммы RP01 и ESY являлись аэробными метилотрофными бактериями,
предпочтительным источником углерода и энергии для которых являлся метанол. Он
использовался в диапазоне концентраций от 0.01 до 3 %, с оптимумом при 0.1-0.5%.
Помимо метанола, новые изоляты обнаруживали слабый рост на этаноле, малате,
валерате и капронате (Рис. 7). Таким образом, спектр утилизируемых штаммами RP01
и ESY субстратов отличался от такового у штаммов BW863T и BW872.
12OD600
0.4
метанол
капронат
0.3
валерат
малат
этанол
0.2
контроль
0.1
0
Сутки
0 5 10 15
Рис 7. Рост штамма ESY на средах с разными источниками углерода.
Штаммы RP01 и ESY являлись ацидофильными организмами, способными к
росту при pH от 4 до 7, с оптимумом при pH 4.7-5.3. Они росли в диапазоне
температур от +4 до +30˚С, с оптимумом при 18-22ºС. Ингибирование роста
происходило при содержании NaCl > 0.7 %. Содержание пар Г+Ц в ДНК - 60.6 моль
%. Уровень ДНК-ДНК гибридизации штаммов BW863T и ESY составлял 35%, что
свидетельствует о принадлежности последнего к новому виду рода Methylovirgula.
Таким образом, можно заключить, что метилотрофные бактерии рода
Methylovirgula населяют кислые наземные экосистемы различных типов и
представляют ранее некультивируемую группу бактерий, важная роль которых в
процессе окисления метанола в данных местообитаниях была установлена
молекулярными методами.
2. Новые факультативно метанотрофные бактерии семейства
Beijerinckiaceae.
2.1. Метанотрофы с растворимой формой ММО.
Из образцов торфа двух сфагновых болот бореальной зоны России - болота
Торфяное, Архангельской обл. и болота Бакчарское, Томской обл. - были получены
стабильные метанотрофные сообщества, из которых были выделены 2 штамма
метанотрофных бактерий – AR4T и SOP9, соответственно. Еще один штамм – LAY –
был выделен P. Dunfield из кислой лесной почвы около Марбурга, Германия. Все три
штамма представляли собой образующие розетки клетки палочковидной формы
размером 1.1-2.9 мкм в длину и 0.4-0.65 мкм в ширину (Рис. 8а). Анализ ультратонких
срезов клеток, выращенных на метане, продемонстрировал отсутствие
внутрицитоплазматических мембран (Рис. 8в), что характерно только для
факультативных метанотрофов рода Methylocella, обладающих растворимой ММО.
Однако морфология клеток новых изолятов была принципиально отлична от таковой
у Methylocella spp. (Рис. 8б).
13Рис. 8. Морфология клеток штамма AR4T (а) по сравнению с Methylocella spp. (б).
Ультратонкое строение клеток штамма AR4T (в). Маркеры – 2 мкм.
5
ppm * 10 OD600
0.1
2.5
0.08
2
0.06
1.5
1 0.04
0.5 0.02
0 0
0 5 10 15 20 25 Сутки 30
Метан Контроль AR4 Контроль
Рис. 9. Рост штамма AR4T на метане в качестве единственного источника углерода и
энергии. Черные круги – рост штамма AR4Т на метане (OD600), черные квадраты –
убыль субстрата (метана), белые круги – рост в контроле без метана, белые квадраты –
динамика концентрации метана в контрольном варианте без бактериальной культуры.
Ближайшими филогенетическими родственниками штаммов LAY, AR4T и SOP9
оказались метанотрофы рода Methylocella и гетеротрофы рода Beijerinckia (96-96.5%
сходства последовательностей гена 16S рРНК) (Рис. 1). Подобно Methylocella spp.,
новые изоляты были способны расти на метане как единственном источнике углерода
и энергии (Рис. 9).
Попытки амплифицировать ген pmoA, кодирующий полипептид, несущий
активный центр мембраной ММО, со всеми ныне разработанными парами праймеров
результата не дали. Подобно представителям Methylocella spp., у новых изолятов
14присутствовала только растворимая форма этого фермента, что было подтверждено
амплификацией гена mmoX, кодирующего α -субчастицу растворимой ММО (Рис. 10).
Среди известных метанотрофов, последовательности mmoX новых изолятов были
наиболее близки таковым у рода Methylocella, но формировали отдельный кластер,
обнаруживая высокое сходство с группой последовательностей mmoX клонов,
полученных ранее из кислых болот различных типов (Chen et al., 2008).
Рис. 10. Дендрограмма, построенная на основе сравнительного анализа нуклеотидных
последовательностей гена mmoX, показывающая положение новых штаммов AR4T,
LAY и SOP9 относительно некоторых метанотрофных представителей Proteobacteria, а
также последовательностей, выявленных в кислых болотах молекулярными методами
(Chen et al., 2008). В качестве внешней группы использованы нуклеотидные
последовательности генов mmoX метанотрофов I типа. Маркер – 0.05 замещений на
нуклеотидную позицию.
Помимо метана, штаммы хорошо росли на метаноле, а также обнаруживали
способность к росту на некоторых органических кислотах: ацетате, малате, пирувате
и сукцинате. Таким образом, спектр субстратов, используемых штаммами AR4T, LAY
и SOP9, был близок к таковому у представителей рода Methylocella. Единственным
отличием явилось отсутствие роста новых изолятов на этаноле.
Штаммы AR4T, LAY и SOP9 являлись мезофильными и ацидотолерантными
организмами, растущими в диапазоне температур от +4 до +33˚С и при pH от 3.5 до
7.2. Оптимальный рост наблюдался при 20-23ºС и при pH 4.8-5.2 (Рис. 11 А, Б).
По сравнению с представителями Methylocella spp., новые изоляты являлись
более ацидотолерантными организмами. Кроме того, они оказались первыми
известными факультативными метанотрофами, способными к росту при pH ниже 4.
Отличия в морфологии клеток, спектре используемых субстратов, ростовом
диапазоне значений pH, составе клеточных жирных кислот (Таблица 1) и
генотипических характеристиках позволили предложить описать штаммы AR4T, LAY
15и SOP9 в качестве нового рода и вида семейства Beijerinckiaceae – ‘Methyloferula
sphagni’ gen. nov., sp. nov.
µ, hr-1 µ, hr-1
А Б
0.016
0.03
0.012
0.02
0.008
0.01 0.004
0 0
2.5 3.5 4.5 5.5 6.5 7.5 0 10 20 30 TºC
pH
Рис. 11. Зависимость удельной скорости роста штамма AR4T от величины pH (А) и
температуры (Б). Для сравнения пунктирной линией и белыми кругами показана
зависимость удельной скорости роста Methylocella spp. от величины pH.
Характеристика ‘Methyloferula’ gen. nov. Methyloferula (methyl от Gr. n. methu,
вино и Gr. n. hulê, дерево, метильная группа; L. fem. n. ferula, палочка; N.L. fem. n.
Methyloferula, метилотрофная палочковидная клетка).
Одиночные грамотрицательные клетки палочковидной формы, образующие
розетки или скопления неправильной формы. Размножаются неравномерным
делением. Колонии выпуклые, непрозрачные, кремово-белые, диаметром 0.3-1.2 мм,
плотной консистенции, с ровным краем и гладкой поверхностью. Рост культур в
жидких средах гомогенный, но в старых культурах наблюдается образование хлопьев.
Аэробные факультативные метанотрофы с растворимой формой ММО.
Предпочтительный субстрат – метанол. Способны использовать ряд органических
кислот: ацетат, малат, пируват, сукцинат. Ацидотолерантные, мезофильные
организмы. Чувствительны к концентрациям NaCl выше 0.7%. Преобладающие
жирные кислоты – 18:1ω7c и cy19:1ω8c. Содержание пар Г+Ц в ДНК 55.6-57.5 моль
%. Род принадлежит к классу Alphaproteobacteria, семейству Beijerinckiaceae.
Типовой вид – Methyloferula sphagni.
‘Methyloferula sphagni’ sp. nov. Methyloferula sphagni (sphag'ni. L. gen. n. sphagni,
из сфагнума, изолирован из сфагнового торфа). Основные характеристики указаны
при описании рода. Размер клеток 1.1-2.9 мкм в длину и 0.4-0.65 мкм ширину.
Источники азота - аммоний, нитрат, дрожжевой экстракт и мочевина. Не нуждаются в
факторах роста. Растут при температуре от 4 до 33˚C и в диапазоне pH 3.5-7.2.
Оптимальный рост при 20-23˚C, и при pH 4.8-5.2. Местообитания – болота и почвы с
низкими значениями pH. Типовой штамм AR4T (ВКМ В-2543T), изолирован из
кислого сфагнового болота Торфяное Архангельской обл. (65º01'N, 35º44'E).
162.2. Новые метанотрофы с мембранной формой ММО.
Еще один изолят факультативно метанотрофной бактерии, штамм KYGT был
выделен P. Dunfield из осадков ручья в лесном массиве близ Марбурга, Германия.
Ближайшим филогенетическим родственником (98% сходства последовательностей
гена 16S рРНК) этого изолята являлся облигатный метанотроф Methylocapsa
acidiphila B2 (Рис. 1). Морфологически, штамм KYGT представлял собой короткие
изогнутые палочки с биполярно расположенными гранулами поли-β-
гидроксибутирата (Рис. 12а, б). Клетки содержали стопки хорошо развитых
внутрицитоплазматических мембран, что характерно и для M. acidiphila (Рис. 12б).
Рис. 12. Морфология и ультратонкое строение клеток штамма KYGT: (a) фазово-
контрастная микрофотография клеток, маркер – 2 мкм; (б) электронная
микрофотография ультратонкого среза клетки, маркер – 1 мкм.
Предпочтительным источником углерода и энергии для штамма KYGT являлся
метан. Однако, в отличие от M. acidiphila, у нового изолята была обнаружена
способность к росту не только на С1-соединениях, метане и метаноле, но также на
ацетате (Рис. 13). Факультативные метанотрофы рода Methylocella, не имеющие
мембранной формы ММО, также способны к росту на ацетате (Dedysh et al., 2005).
Однако, в отличие от представителей Methylocella spp., которые предпочитают расти
на ацетате, штамм KYGT гораздо лучше рос на метане (ODmax=1.2, µ=0.018 час-1), чем
на ацетате (ODmax=0.3, µ=0.006 час-1). Новый изолят обладал только мембранной
формой ММО. Сравнительный анализ гена pmoA показал, что последовательности
PmoA штамма KYGT наиболее близки транслированным аминокислотным
последовательностям pmoA Methylocapsa аcidiphila B2 (Рис. 14).
На основании анализа морфологических, хемотаксономических и
генотипических характеристик было предложено отнести новый изолят к новому
виду рода Methylocapsa – ‘Methylocapsa aurea’ sp. nov.
Характеристика Methylocapsa aurea sp nov. Methylocapsa aurea (au’re.a. L. fem.
adj. aurea золотой, желтый цвет колоний). Грамотрицательные палочковидные клетки
17OD600
0.8
0.6
0.4
0.2
0
0 2 4 6 8 10 12 14 16 Сутки
ацетат метан контроль
Рис. 13. Рост штамма KYGT на средах с разными источниками углерода.
Рис. 14. Дендрограмма, построенная на основе сравнительного анализа
транслированных аминокислотных последовательностей гена pmoA штамма KYGT и
других метанотрофов. Маркер, 0.1 замещений на нуклеотидную позицию.
размером 0.7-1.2 мкм на 1.8-3.1 мкм, содержащие стопки ВЦМ и биполярно
расположенные гранулы поли-β-гидроксибутирата. Колонии желтоватого цвета.
Используют метан, метанол и ацетат. Источника азота - аммоний, нитрат, мочевина,
L-пролин, L-аланин, L-аргинин, пептон и дрожжевой экстракт. Фиксируют
молекулярный азот посредством кислород-чувствительной нитрогеназы.
Оптимальный рост при значениях температуры 25-30ºC и pH 6.0-6.2. Рост
ингибируется при содержании NaCl в среде выше 0.2-0.3%. Преобладающая жирная
кислота – 18:1ω7c. Содержание пар Г+Ц в ДНК 60.2 моль %. Типовой штамм KYGT
(=DSM 22158T =VKM B-2544T), выделен из кислой лесной почвы окрестностей
Марбурга, Германия.
183. Оценка роли факультативных метанотрофов в общей структуре
метанокисляющего фильтра болотных экосистем.
3.1 Численность факультативных метанотрофов родов Methylocella и
‘Methyloferula’ в сфагновом торфе.
Для целей детекции in situ новых факультативных метанотрофов рода
‘Methyloferula’ был разработан зонд Mfer-431 (5-TTCCCGGGCAAAAGAGCT-3),
который был применен для определения численности целевой популяции в образцах
торфа, отобранных из слоя 0-10 см болота Бакчарское, Томской области. Новый зонд
использовали в сочетании с ранее предложенными для метанотрофов
олигонуклеотидными зондами разного уровня специфичности (см. Материалы и
методы). Общая численность выявленных в образцах метанотрофных бактерий
варьировала от 4.6×106 до 1.4×107 клеток г-1 торфа, составляя до 15% общего числа
клеток бактерий. Численность метанотрофов I типа (гибридизация с зондами М84 +
М705) была незначительной (0.3 – 6.3×105 клеток г-1 торфа). Обнаруженные в торфе
метанотрофы II типа были представлены двумя группами: Methylosinus/Methylocystis
(детекция зондом М-450) и Methylocella/Methylocapsa (зонды Mcell-1026/Mcaps-1032).
В составе сообщества доминировали представители группы
Methylosinus/Methylocystis, составляя до 76% клеток всех метанотрофов. Численность
клеток рода Methylocapsa составляла около 1% общего числа метанотрофов.
Применение нового зонда Mfer-431 показало, что численность ‘Methyloferula sphagni’
достигает 3.5×106 клеток г-1 торфа. Таким образом, доля факультативных
метанотрофов родов Methylocella и ‘Methyloferula’ составляла 20-25% всех
метанотрофных бактерий, выявленных в торфе методом FISH (Рис. 14).
Рис. 14. Соотношение численности клеток отдельных групп метанотрофных бактерий,
выявленных методом FISH в образцах сфагнового торфа болота Бакчарское.
193.2 Вклад факультативных метанотрофов в общую метанокисляющую
активность сфагнового торфа.
Особенность регуляции окисления метана у факультативных метанотрофов с
растворимой ММО (Methylocella и ‘Methyloferula’) состоит в ингибировании
активности ММО при наличии в среде органических субстратов роста, например,
ацетата (Dedysh et al., 2005; Theisen et al., 2005). Это дает возможность оценить вклад
этих факультативных метанотрофов в общую активность окисления метана в
сфагновом торфе путем внесения ацетата. Результаты такого модельного
эксперимента показаны на Рис. 15. Как видно на графике, внесение ацетата в образец
сфагнового торфа до концентрации 0.5 мМ вызывало снижение общей скорости
окисления метана в 2 раза, свидетельствуя о переключении факультативных
метанотрофов с метанотрофного типа метаболизма на гетеротрофный. Таким
образом, вклад факультативных метанотрофов с растворимой формой ММО в общую
активность окисления метана в кислых экосистемах может быть весьма
значительным. Возможно также, что разнообразие этих организмов не исчерпывается
представителями родов Methylocella и ‘Methyloferula’, и существуют бактерии с
аналогичным типом метаболизма, которые не выявляются ныне имеющимися
олигонуклеотидными зондами.
Ацетат
Рис. 15. Влияние внесения ацетата (показано стрелкой) на активность окисления СН4 в
образцах сфагнового торфа. Черные круги – динамика убыли метана в образцах торфа
без ацетата, белые круги – динамика убыли метана в образцах после внесения ацетата.
20ЗАКЛЮЧЕНИЕ
Настоящее исследование значительно расширило разнообразие известных
представителей семейства Beijerinckiaceae и показало, что эти бактерии являются
функционально-значимым компонентом микробного сообщества кислых наземных
экосистем. Уникальность этого семейства состоит в том, что оно объединяет
филогенетически близкородственные организмы (с уровнем сходства генов 16S рРНК
96% и выше), обладающие различными типами метаболизма – от типично
гетеротрофного до облигатно метанотрофного (Таблица 2). Это обстоятельство
делает невозможным установление способности микроорганизма из Beijerinckiaceae к
использованию С1-соединений на основании одних только данных анализа генов 16S
рРНК и требует обязательной идентификации функциональных генов, кодирующих
ММО или МДГ.
Выделенные и охарактеризованные в настоящем исследовании микроорганизмы
– два новых рода и три новых вида – представляют собой метано- и метилотрофные
бактерии, ближайшими филогенетическими родственниками которых являются
метанотрофы родов Methylocapsa и Methylocella, а также гетеротрофы рода
Bejerinckia. Эти бактерии хорошо адаптированы к существованию при низких
значениях pH, характерных для большинства наземных экосистем Северного
полушария. Представители вновь описанного рода ‘Methyloferula’ обладают одной
только растворимой формой ММО, а предпочтительными субстратами этих
организмов являются метанол и ряд органических кислот. Наличие последних в среде
ингибирует окисление метана. Другая группа метанотрофов семейства
Beijerinckiaceae, представленная родом Methylocapsa, характеризуются обладанием
только мембранной формой ММО и предпочтением метана в качестве ростового
субстрата. Тем не менее, в отсутствие метана эти метанотрофы способны к
медленному росту на ацетате. Таким образом, разнообразие метаболических
подтипов метанотрофных бактерий в природе оказалось шире такового,
представленного ранее в культурах коллекций. Вклад облигатно- и факультативно
метанотрофных бактерий в суммарную активность окисления метана в различных
экосистемах еще предстоит оценить, но настоящее исследование показывает, что
вклад факультативных метанотрофов может быть весьма существенен в
местообитаниях с кислой реакцией среды.
21Таблица 2. Дифференцирующие характеристики представителей семейства
Beijerinckiaceae.
Характеристи Beijerinckia Methylovir-
‘Methyloferula’ Methylocapsa Methylocella
ка gula
Биполярные Прямые
Прямые или Изогнутые Биполярные
Морфология прямые или или
изогнутые кокковидные прямые или
клеток изогнутые изогнутые
палочки клетки изогнутые
палочки палочки
палочки
0.5 – 1.5 × 0.4 – 0.65 × 0.4 – 0.65 × 0.7 – 1.2 × 0.6 – 1.0 ×
Размер клеток,
1.7 – 4.5 1.2 – 2.5 1.1 – 2.9 0.8 – 3.1 1.0 – 2.5
мкм
Образование _
+ + – –
розеток
Облигатная
Факультати
Гетеротрофия или
Тип вная Факультативная Факультативная
или факультативн
метаболизма метилотроф метанотрофия метанотрофия
метилотрофия ая
ия
метанотрофия
_
Рост на метане – + + +
Форма ММО _ _ рММО мММО рММО
Сахара, орг. Этанол, Ацетат, Ацетат,
Используемые
кислоты, пируват, пируват, Нет или пируват,
органические
спирты сукцинат, сукцинат, ацетат сукцинат,
субстраты
малат малат малат, этанол
Потребность в _
+ – – –
факторах роста
Рост при:
35ºC + – + – –
pH 7.0 + – + + +
pH 3.5 + + + – –
0.5% NaCl – + – – –
Содержание
G+C (мол%) 54.7-59.1 61.8-62.8 55.6-57.5 57.3 60-63.3
22ВЫВОДЫ
1. Расширено таксономически охарактеризованное разнообразие метано- и
метилотрофных представителей семейства Beijerinckiaceae, включающего
филогенетически близкие организмы с различными типами метаболизма. В отличие
от семейств Methylococcaceae и Methylocystaceae, установление способности
микроорганизма из Beijerinckiaceae к использованию С1-соединений невозможно на
основании одних только данных анализа генов 16S рРНК.
2. Выделенные из кислой почвы штаммы облигатно ацидофильных
метилотрофных бактерий описаны в качестве нового рода и вида Methylovirgula ligni
gen. nov., sp. nov. Они являются первыми представителями выявленной ранее
молекулярными методами некультивируемой группы метилотрофных бактерий,
осуществляющих окисление метанола в кислых почвах.
3. Три штамма факультативно метанотрофных бактерий с растворимой формой
ММО, изолированные из сфагновых болот и кислой почвы, предложено описать в
качестве нового рода и вида ‘Methyloferula sphagni’ gen. nov., sp. nov. Эти организмы
являются единственными ныне известными факультативными метанотрофами,
способными к росту при значениях pH ниже 4.
4. Один штамм факультативно метанотрофных бактерий с мембранной формой
ММО предложено описать в качестве нового вида рода Methylocapsa, ‘Methylocapsa
aurea’ sp. nov. Впервые показана способность использования ацетата в качестве
субстрата роста представителями рода Methylocapsa, ранее считавшимися
облигатными метанотрофами.
5. Выявлена высокая численность и установлен значительный вклад
факультативных метанотрофов с растворимой формой ММО в общую активность
окисления метана в кислых сфагновых болотах.
23Список работ, опубликованных по теме диссертации
Экспериментальные статьи
1. Воробьев А.В., Дедыш С.Н. Использование накопительных культур для оценки
структуры сообществ метанотрофов: проблема репрезентативности результатов.
Микробиология, 2008, Т. 77, № 4, С. 566–569.
2. Vorob’ev A.V., de Boer W., Folman L.B., Bodelier P.L.E., Doronina N.V., Suzina N.E.,
Trotsenko Y.A., Dedysh S.N. Methylovirgula ligni gen. nov., sp. nov., an obligately
acidophilic, facultatively methylotrophic bacterium with highly divergent mxaF gene. Int J
Syst Evol Microbiol, 2009, V. 59 (10), p. 2538 - 2545.
3. Dunfield P.F., Belova S.E., Vorob’ev A.V., Cornish S.L., Dedysh S.N. Methylocapsa
aurea sp. nov., a facultatively methanotrophic bacterium possessing a particulate methane
monooxygenase. Int J Syst Evol Microbiol (submitted).
Тезисы
1. Воробьев А.В., Дедыш С.Н. Эмиссия метана и численность метанотрофных
бактерий в различных локусах сфагнового болота: влияние доминирующих
растительных ассоциаций. Материалы II Международной молодежной школы-
конференции «Актуальные аспекты современной микробиологии». Москва, ИНМИ
РАН. 1-3.11.2007. МАКС Пресс, Москва. С. 58-60.
2. Vorob’ev A.V., de Boer W., Dedysh S.N. Characterization of novel acidophilic
methylotrophic bacteria with highly divergent mxaF genes. 12th International Symposium on
Microbial Ecology. August 17-22, 2008, Cairns, Australia. Abstract book, p.35.
3. Vorob’ev A.V., Glagolev M.V., Dedysh S.N. Methane fluxes and methanotroph
community structures in wetland sites with different vegetation covers. 12th International
Symposium on Microbial Ecology. August 17-22, 2008, Cairns, Australia. Abstract book,
p.202.
4. Воробьев А.В. Новые метило- и метанотрофные бактерии из кислых наземных
экосистем бореальной зоны. Тезисы докладов XIII Международной конференции
студентов и аспирантов по фундаментальным наукам «Ломоносов 2009», Секция
Биология». М. МГУ. 2009.
5. Vorob’ev A.V., Dedysh S.N. Novel facultatively methanotrophic bacteria from acidic
sphagnum weatlands. 3th Congress of European microbiologists. June 28 – July 2, 2009,
Gothenburg, Sweden. Abstract book, p.183.
24Вы также можете почитать

















































